M1-Linked Ubiquitin Chains: Decoding the Linear Signal in Immune Signaling and Therapeutic Intervention
This article provides a comprehensive exploration of the functions of M1-linked (linear) ubiquitin chains in immune signaling, tailored for researchers and drug development professionals.
M1-Linked Ubiquitin Chains: Decoding the Linear Signal in Immune Signaling and Therapeutic Intervention
Abstract
This article provides a comprehensive exploration of the functions of M1-linked (linear) ubiquitin chains in immune signaling, tailored for researchers and drug development professionals. It covers the foundational biology of the Linear Ubiquitin Chain Assembly Complex (LUBAC) and its role in regulating NF-κB activation, cell survival, and inflammatory responses. The scope extends to current methodologies for detecting and quantifying M1-linked ubiquitination, addresses common experimental challenges, and offers a comparative analysis with other ubiquitin linkage types. By synthesizing recent advances, this review highlights the significant potential of targeting the linear ubiquitination machinery for treating inflammatory diseases, cancer, and immune disorders.
The Linear Ubiquitin Code: Assembly, Architecture, and Core Signaling Functions in Immunity
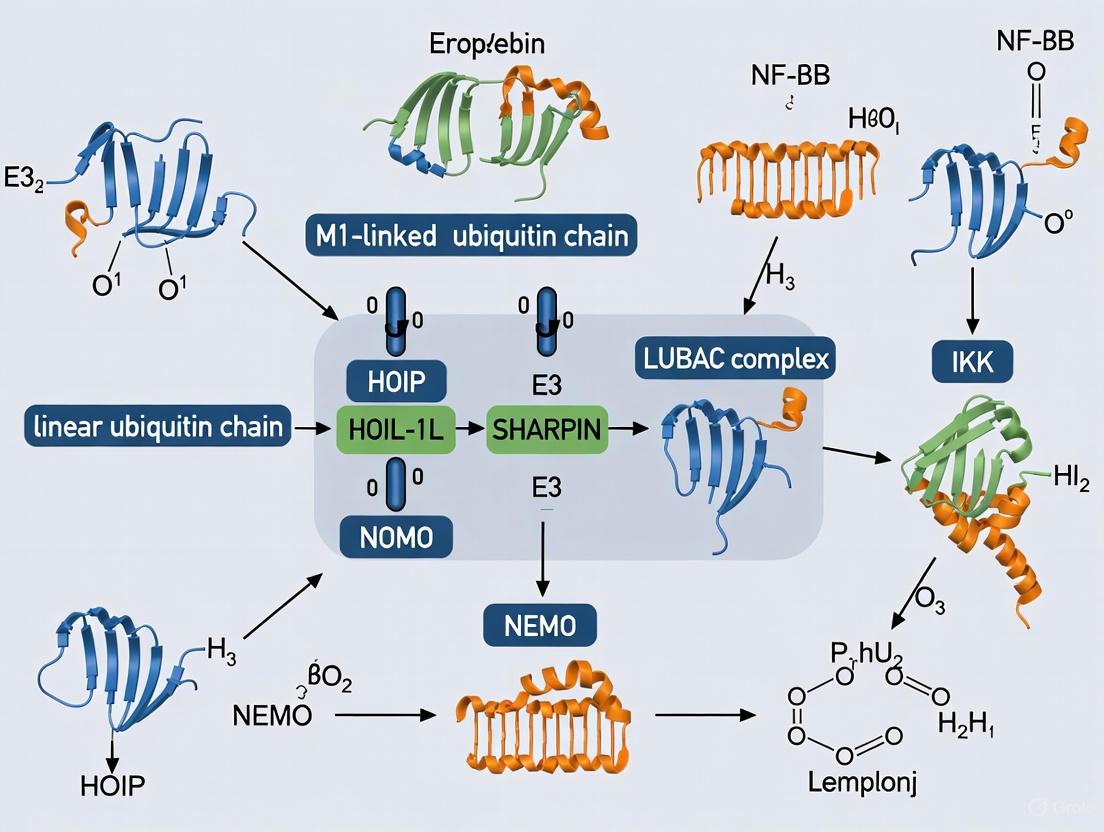
The Linear Ubiquitin Chain Assembly Complex (LUBAC) represents a paradigm shift in ubiquitin research as the only identified E3 ubiquitin ligase complex that specifically catalyzes the formation of M1-linked linear ubiquitin chains, distinct from the more common lysine-linked ubiquitin chains [1]. Discovered as a ~600 kDa complex, LUBAC functions as a critical signaling hub in immune regulation, cell death pathways, and inflammation, making it a focal point for therapeutic intervention in autoimmune diseases, inflammatory conditions, and cancer [1] [2]. The complex consists of three core subunits: the catalytic component HOIP (RNF31), and two accessory subunits HOIL-1L (RBCK1) and SHARPIN, which form a tight 1:1:1 stoichiometric complex [3]. Through its unique ability to generate linear ubiquitin chains, LUBAC modulates key signaling pathways including NF-κB activation, TNF signaling, and various cell death pathways (apoptosis, necroptosis, and pyroptosis) by providing specific scaffolding platforms for the assembly of signaling complexes [4] [1]. The precise structural organization and mechanistic regulation of LUBAC components determine its cellular functions and have become targets for drug development in various human diseases.
Structural Organization of LUBAC Components
HOIP: The Catalytic Core of LUBAC
HOIP serves as the catalytically essential component of LUBAC, containing several critical domains that regulate its E3 ligase activity and interaction with other complex components. The RING-In-Between-RING (RBR) domain represents the catalytic heart of HOIP, employing a unique RING/HECT hybrid mechanism for ubiquitin transfer [5]. Unlike typical RING ligases that directly transfer ubiquitin from E2 to substrate, HOIP forms a transient thioester intermediate with ubiquitin on its Cys885 residue within the RING2 domain before transferring it to the substrate [1] [2]. The Linear ubiquitin chain Determining Domain (LDD) at the C-terminus is unique to HOIP and is responsible for specifically recognizing the acceptor ubiquitin and positioning its N-terminal methionine for linear chain formation [1] [5]. The Ubiquitin-Associated (UBA) domain of HOIP interacts with the ubiquitin-like (UBL) domains of both HOIL-1L and SHARPIN, facilitating complex assembly [6] [7]. Importantly, HOIP's catalytic activity is autoinhibited by its N-terminal domain, and this inhibition is relieved upon binding of HOIL-1L and SHARPIN, which induces conformational changes that activate the enzyme [5].
Table 1: Key Structural Domains of HOIP and Their Functions
| Domain | Location | Structural Features | Functional Role |
|---|---|---|---|
| UBA Domain | N-terminal | Binds UBL domains | Recruits HOIL-1L and SHARPIN; relieves autoinhibition |
| RING1 Domain | Central | Zinc-binding motif | Recognizes and binds E2~Ub conjugate (UBE2L3) |
| IBR Domain | Central | In-Between-RING motif | Connects RING1 and RING2 domains |
| RING2 Domain | Central | Catalytic cysteine (Cys885) | Forms thioester with ubiquitin; catalytic intermediate |
| LDD Domain | C-terminal | Unique to HOIP | Recognizes acceptor ubiquitin; determines linear linkage specificity |
HOIL-1L and SHARPIN: Accessory Subunits with Regulatory Functions
HOIL-1L and SHARPIN play crucial roles in stabilizing the LUBAC complex and regulating its catalytic activity through multiple interconnected mechanisms. Both proteins contain Ubiquitin-Like (UBL) domains that interact with the UBA domain of HOIP, facilitating their incorporation into the complex [6] [7]. The LUBAC-Tethering Motif (LTM) domains, located N-terminally to the UBL domains in both proteins, heterodimerize and fold into a single globular domain that is critical for stabilizing the trimeric LUBAC complex [7] [5]. This LTM-mediated interaction is remarkably resistant to dissociation and plays an indispensable role in maintaining complex integrity [7]. Additionally, both HOIL-1L and SHARPIN contain Npl4 Zinc Finger (NZF) domains that facilitate ubiquitin binding, though with different specificities: HOIL-1L NZF displays robust binding affinity for linear ubiquitin chains, while SHARPIN NZF binds both linear and K63-linked ubiquitin chains [8]. HOIL-1L itself is an RBR-type E3 ligase with catalytic activity, capable of forming oxyester bonds between ubiquitin and serine/threonine residues of substrates, and it monoubiquitinates LUBAC subunits to regulate complex activity [3] [2].
Table 2: Comparative Analysis of HOIL-1L and SHARPIN Domains
| Feature | HOIL-1L | SHARPIN |
|---|---|---|
| Molecular Weight | ~59 kDa | ~43 kDa |
| Domain Organization | LTM-UBL-NZF-RBR | LTM-UBL-NZF |
| LTM Function | Heterodimerization with SHARPIN LTM | Heterodimerization with HOIL-1L LTM |
| UBL Function | Binds HOIP UBA domain | Binds HOIP UBA domain |
| NZF Specificity | High affinity for linear ubiquitin chains | Binds linear and K63-linked ubiquitin chains |
| Catalytic Activity | RBR domain with E3 activity (oxyester bonds) | Non-catalytic |
| Stabilization Role | Critical for LUBAC integrity | Critical for LUBAC integrity |
Structural Architecture of the Holocomplex
The three-dimensional architecture of LUBAC reveals an elongated asymmetric crescent structure with the majority of the mass concentrated at one end, as determined by electron microscopy and crystal structure analyses [3]. The complex exhibits a 1:1:1 stoichiometry of HOIP:HOIL-1L:SHARPIN, though it can form higher-order oligomers under certain conditions [3]. The interaction between the UBL domains of HOIL-1L/SHARPIN and the UBA domain of HOIP induces allosteric rearrangements that reorient the UBA and RBR-LDD domains, facilitating E2 loading and promoting the catalytic activity of HOIP [6] [5]. The coordinated ubiquitin relay mechanism between HOIP and HOIL-1L ligase activities enables the formation of heterotypic ubiquitin chains containing linear linkages with oxyester-linked branches, expanding the regulatory potential of LUBAC-mediated ubiquitination [3].
Molecular Mechanisms of LUBAC Activation and Regulation
Allosteric Activation of HOIP Catalytic Activity
The catalytic core of LUBAC is tightly regulated through sophisticated allosteric mechanisms that ensure precise temporal and spatial control of linear ubiquitination. In its basal state, full-length HOIP exhibits autoinhibition through intramolecular interactions involving its N-terminal domain, which suppresses catalytic activity in the absence of accessory subunits [5]. Binding of the UBL domains of HOIL-1L and SHARPIN to the UBA domain of HOIP releases this autoinhibition by inducing conformational changes that reorient the UBA domain relative to the RBR-LDD region [6] [5]. Structural studies have demonstrated that HOIL-1L and SHARPIN can separately or synergistically bind to distinct sites within the HOIP UBA domain, with their combined binding inducing optimal allosteric effects that facilitate E2 loading and enhance catalytic efficiency [6]. This allosteric rearrangement properly positions the catalytic Cys885 residue in the RING2 domain for thioester bond formation with ubiquitin and enables the LDD domain to specifically recognize the acceptor ubiquitin and facilitate Met1 linkage formation [1] [5].
Coordination Between HOIP and HOIL-1L Catalytic Activities
The concerted actions of HOIP and HOIL-1L catalytic activities enable LUBAC to generate complex heterotypic ubiquitin chains with diverse signaling capabilities. HOIL-1L functions as an atypical E3 ligase that catalyzes the formation of oxyester bonds between the C-terminus of ubiquitin and serine/threonine residues of substrates, in contrast to the isopeptide bonds formed by conventional E3 ligases [3]. Recent research has revealed that HOIL-1L-mediated oxyester ubiquitination occurs within the LUBAC complex and can generate heterotypic ubiquitin chains containing linear linkages with oxyester-linked branches [3]. Cross-linking mass spectrometry studies indicate proximity between the RBR domains of HOIP and HOIL-1L, suggesting a coordinated ubiquitin relay mechanism between the two ligases [3]. Additionally, HOIL-1L monoubiquitinates all three LUBAC subunits via isopeptide linkages to lysine residues, which subsequently serves as a priming step for HOIP-mediated conjugation of linear chains onto the monoubiquitin, creating an auto-regulatory mechanism that can attenuate LUBAC function [1] [2].
Ubiquitin Binding and Recruitment Mechanisms
The NZF domains of HOIL-1L and SHARPIN play complementary roles in recruiting LUBAC to specific signaling complexes and regulating its function through ubiquitin binding. HOIL-1L NZF displays robust and specific binding to linear ubiquitin chains, while SHARPIN NZF exhibits dual specificity, binding both linear and K63-linked ubiquitin chains with moderate affinity [8]. This synergistic ubiquitin-binding capability allows LUBAC to be recruited to signaling complexes containing different ubiquitin chain types and to amplify linear ubiquitination in specific subcellular locations. Experimental evidence demonstrates that the ubiquitin-binding activities of both NZF domains cooperatively regulate LUBAC functions, with HOIL-1L NZF playing a predominant role in NF-κB activation, while SHARPIN NZF primarily contributes to cell death protection [8]. Mutations that disrupt the ubiquitin-binding capacity of both NZF domains simultaneously profoundly impair both NF-κB activation and cell death protection functions, highlighting their critical and synergistic importance in LUBAC regulation [8].
Methodologies for Studying LUBAC Structure and Function
Structural Biology Approaches
Elucidating the three-dimensional architecture of LUBAC has required the integration of multiple structural biology techniques, each providing complementary insights into complex organization and dynamics. X-ray crystallography of core domains has revealed atomic-level details of key interactions, particularly the co-crystal structure of the binding regions demonstrating that LTMs of HOIL-1L and SHARPIN heterodimerize to form a single globular domain critical for trimeric LUBAC stabilization [7]. Electron microscopy (EM) has provided the first 3D reconstruction of the full LUBAC holoenzyme, revealing an elongated asymmetric crescent structure with majority mass concentration at one end and confirming a monodisperse distribution of particles with distinct dumbbell morphology [3]. Cross-linking mass spectrometry has been instrumental in mapping proximities between catalytic RBR domains of HOIP and HOIL-1L, supporting a coordinated ubiquitin relay mechanism between the two ligases and identifying interaction interfaces difficult to capture by other methods [3]. Nuclear Magnetic Resonance (NMR) spectroscopy has characterized domain dynamics and binding interactions, particularly for the NZF domains and their ubiquitin-binding specificities, revealing differential affinities for various ubiquitin chain types [8].
Biochemical and Biophysical Assays
A comprehensive suite of biochemical and biophysical assays has been developed to quantify LUBAC activity, interactions, and stability. Size exclusion chromatography with multi-angle light scattering (SEC-MALS) has confirmed the oligomeric state and stoichiometry of LUBAC components, identifying multiple populations with different oligomeric states and verifying the 1:1:1 stoichiometry of the core complex [3]. Isothermal titration calorimetry (ITC) and surface plasmon resonance (SPR) have precisely quantified binding affinities between components, particularly the UBA-UBL interactions and NZF-ubiquitin interactions, revealing synergistic binding effects [6] [8]. In vitro ubiquitination assays using purified components have delineated the individual contributions of each subunit, demonstrating that HOIL-1L and SHARPIN synergistically activate HOIP catalysis and facilitate E2 loading [6] [5]. Ubiquitin binding assays using purified NZF domains and different ubiquitin chain types have characterized binding specificities, confirming HOIL-1L NZF preference for linear chains while SHARPIN NZF binds both linear and K63-linked chains [8].
Table 3: Key Experimental Methods for LUBAC Characterization
| Method Category | Specific Techniques | Key Applications | Notable Findings |
|---|---|---|---|
| Structural Biology | X-ray crystallography, Cryo-EM, NMR, Cross-linking MS | 3D structure determination, Domain interactions, Complex architecture | LTM heterodimerization, Allosteric activation mechanism, 3D reconstruction of holoenzyme |
| Biochemical Assays | SEC-MALS, ITC, SPR, In vitro ubiquitination | Binding affinity, Stoichiometry, Catalytic activity, Chain specificity | Synergistic UBA-UBL binding, E2 loading facilitation, Heterotypic chain formation |
| Cellular Studies | CRISPR/Cas9 KO, Co-immunoprecipitation, Immunoblotting | Cellular functions, Pathway analysis, Complex recruitment | NZF domain cooperation, Complex I recruitment, Cell death protection mechanisms |
| Mutational Analysis | Site-directed mutagenesis, Domain swapping, TF-AA mutants | Functional domain mapping, Ubiquitin binding specificity | Critical residues for ubiquitin binding, Domain functional requirements |
Cellular and Genetic Approaches
Genetic manipulation in cell lines and animal models has been essential for validating structural findings and establishing physiological relevance. CRISPR/Cas9-mediated knockout in mouse embryonic fibroblasts (MEFs) has revealed subunit interdependencies, with HOIP destabilization occurring in the absence of either HOIL-1L or SHARPIN, and has enabled functional studies of NZF domain mutants in NF-κB activation and cell death protection [8]. Reconstitution assays in deficient cell lines using wild-type and mutant constructs have defined functional domains and critical residues, particularly identifying Thr and Phe residues in NZF domains as essential for ubiquitin binding and elucidating cooperative effects between HOIL-1L and SHARPIN NZF domains [8]. Immunoprecipitation and proximity ligation assays have characterized endogenous protein interactions and complex formation, demonstrating LUBAC recruitment to signaling complexes such as TNFR1 Complex I and quantifying changes in recruitment efficiency with NZF domain mutations [8]. Image-based assays for NF-κB translocation and cell death quantification have correlated structural features with functional outputs, using high-content imaging to measure p65 nuclear accumulation and cell viability assays to quantify protection from TNFα-induced apoptosis [9] [8].
The Scientist's Toolkit: Essential Research Reagents and Methodologies
Table 4: Key Research Reagents for LUBAC Investigation
| Reagent Category | Specific Examples | Applications | Technical Considerations |
|---|---|---|---|
| Expression Constructs | Full-length HOIP, HOIL-1L, SHARPIN; Domain truncations; TF-AA NZF mutants | Recombinant protein production, Functional domain mapping, Structure-function studies | Co-expression required for stability; Insect cell system optimal for holoenzyme |
| Cell Lines | HOIP/HOIL-1L/SHARPIN knockout MEFs; Jurkat T cells; Primary human CD4+ T cells | Genetic requirement studies, Signaling pathway analysis, Functional complementation | Partial ablation common in primary cells; consider redundancy and compensation |
| Antibodies | Anti-linear ubiquitin (1E3), Anti-HOIP, Anti-HOIL-1L, Anti-SHARPIN, Phospho-IκBα | Immunoblotting, Immunoprecipitation, Immunofluorescence, Complex isolation | Linear ubiquitin specificity crucial; validate with knockout controls |
| Activity Assays | In vitro ubiquitination with UBE2L3, Ubiquitin binding assays, NF-κB luciferase reporter | Catalytic activity measurement, Ubiquitin chain specificity, Pathway activation | Include autoinhibition controls; optimize E2 concentration and reaction time |
| Chemical Tools | LUBAC inhibitors, TLR signaling inducers (TNFα, LPS), Proteasome inhibitors (MG132) | Pathway modulation, Therapeutic targeting, Mechanism investigation | Specificity validation essential; monitor off-target effects on other ubiquitin pathways |
Implications for Immune Signaling and Therapeutic Applications
Role in NF-κB and Immune Signaling Pathways
LUBAC serves as a critical regulator in multiple immune signaling pathways, with its structural features directly determining its functional specificity in different contexts. In TNFα signaling, LUBAC is recruited to TNF Receptor 1 Complex I through recognition of K63-linked ubiquitin chains by SHARPIN NZF, where it then conjugates linear ubiquitin chains to components including RIPK1 and NEMO, leading to NF-κB activation and suppression of cell death [8]. In T cell receptor (TCR) signaling, LUBAC modulates CBM complex functions downstream of TRAF6, with recent evidence showing that LUBAC acts downstream of TRAF6 to catalyze BCL10 ubiquitination and modulate MALT1 substrate recognition rather than serving as the primary driver of NF-κB activation [9]. The structural organization of LUBAC allows it to function as a specific ubiquitin code writer that integrates signals from different pathways, with the cooperative actions of its subunits ensuring appropriate signaling output based on cellular context. Dysregulation of LUBAC components leads to severe immune disorders, as evidenced by Sharpin-deficient mice developing chronic proliferative dermatitis, and humans with HOIL-1L or HOIP mutations presenting with multiorgan autoinflammatory diseases and immunodeficiency [4] [1].
Therapeutic Targeting and Drug Discovery
The precise structural insights into LUBAC organization have revealed multiple potential targets for therapeutic intervention in cancer, autoimmune, and inflammatory diseases. Small molecule inhibitors targeting the LUBAC-tethering motif interface between HOIL-1L and SHARPIN have shown promise in disrupting complex stability and function, with potential applications in cancers characterized by hyperactive LUBAC signaling [7]. NZF-ubiquitin interaction inhibitors have been identified through compound screening, with one small molecule demonstrating inhibition of both HOIL-1L and SHARPIN NZF binding to linear ubiquitin chains, effectively suppressing LUBAC-mediated NF-κB activation and cell death protection [8]. PROTAC-based degradation approaches leveraging the ubiquitin-proteasome system to selectively target LUBAC components offer potential for complete pathway ablation in therapeutic contexts where partial inhibition is insufficient [2]. The allosteric regulation of HOIP provides additional targeting opportunities, with potential compounds that stabilize the autoinhibited conformation or prevent accessory subunit binding offering alternative strategies for selective LUBAC inhibition without complete complex disruption [5] [2].
The structural characterization of LUBAC has revealed a sophisticated molecular machine whose precise organization underlies its specific biological functions in immune regulation and cell death control. The intricate domain architecture, allosteric regulation, and synergistic subunit interactions provide multiple layers of control that ensure appropriate linear ubiquitination in response to specific cellular signals. Future research directions include elucidating the structural basis for LUBAC recruitment to specific signaling complexes, determining the molecular mechanisms governing heterotypic ubiquitin chain formation, and developing more specific therapeutic agents that can selectively modulate LUBAC functions in disease contexts. The continuing integration of structural biology, biochemical analysis, and cellular studies will undoubtedly yield further insights into this unique ubiquitin ligase complex and its applications in therapeutic development for immune disorders and cancer.
Mechanisms of M1-Linked Chain Assembly and Deubiquitination by OTULIN and CYLD
M1-linked (linear) ubiquitination is a crucial post-translational modification that regulates inflammatory signaling, cell death, and immunity. This whitepaper details the molecular mechanisms governing the assembly and disassembly of M1-linked ubiquitin chains, with focused examination of the Linear Ubiquitin Chain Assembly Complex (LUBAC) and the deubiquitinases OTULIN and CYLD. We present comprehensive structural and functional analyses, quantitative biochemical data, and detailed experimental methodologies that define the precise regulatory networks controlling linear ubiquitination. Within the broader context of M1 linear ubiquitin chain function in immune signaling research, this guide synthesizes current understanding of how disruption of these mechanisms contributes to inflammatory disease and immune dysregulation, providing researchers and drug development professionals with technical foundations for therapeutic intervention strategies.
Linear ubiquitination constitutes a specialized form of protein ubiquitination where the C-terminal glycine of a distal ubiquitin molecule forms an isopeptide bond with the N-terminal methionine (M1) of a proximal ubiquitin, creating unique polyubiquitin architectures with distinct signaling properties [4] [10]. Unlike canonical ubiquitin linkages that primarily target substrates for proteasomal degradation, M1-linked ubiquitin chains serve as critical scaffolds that facilitate protein-protein interactions in innate immune signaling pathways, including NF-κB activation, MAPK signaling, and the regulation of cell death processes such as apoptosis and necroptosis [4] [11].
The human genome encodes approximately 100 deubiquitinases (DUBs), but only two—OTULIN and CYLD—possess significant activity against M1-linked ubiquitin chains [4] [12]. These DUBs exhibit distinct mechanisms of substrate recognition, chain specificity, and cellular regulation despite targeting the same linkage type. OTULIN stands as the only known mammalian DUB with exclusive specificity for Met1-linked polyubiquitin, while CYLD demonstrates dual specificity for both M1- and K63-linked chains [13] [11]. This review systematically examines the assembly of M1 chains by LUBAC and their disassembly by OTULIN and CYLD, integrating structural insights, quantitative biochemical data, and experimental approaches that define this dynamic regulatory system.
M1-Linked Ubiquitin Chain Assembly by LUBAC
LUBAC Composition and Structure
The Linear Ubiquitin Chain Assembly Complex (LUBAC) represents the sole E3 ubiquitin ligase known to generate M1-linked ubiquitin chains in humans [4]. This multisubunit complex comprises three core components: HOIP (RNF31), HOIL-1L (RBCK1), and SHARPIN, which together form a stable catalytic complex [4]. HOIP serves as the catalytic core of LUBAC, belonging to the RBR (RING-Between-RING-RING) family of E3 ligases and containing a dedicated linear ubiquitin determining domain (LDD) that specifically recognizes acceptor ubiquitin molecules and orients them for M1-linkage formation [4]. HOIL-1L and SHARPIN function as essential regulatory subunits that stabilize the complex through interactions between their ubiquitin-associated (UBA) domains and the N-terminal PUB domain of HOIP [4].
The structural organization of LUBAC components enables precise coordination of the linear ubiquitination process:
- HOIP: Contains RING1, IBR, RING2 domains, and the C-terminal LDD domain that confers specificity for linear chain formation
- HOIL-1L: Features a RBR domain and UBA domain that interacts with HOIP
- SHARPIN: Comprises a PH domain and UBA domain that facilitates HOIP binding
This tripartite architecture allows LUBAC to recognize specific substrates and conjugate M1-linked ubiquitin chains that serve as platforms for signaling assembly in inflammatory pathways [4].
Mechanism of Linear Ubiquitin Chain Formation
LUBAC catalyzes linear ubiquitin chain formation through a multi-step enzymatic process that begins with the recruitment of ubiquitin-charged E2 enzymes. The catalytic mechanism proceeds through several distinct phases:
- E2 Recruitment: HOIP's RING1 domain recruits ubiquitin-charged E2 enzymes (primarily UBE2L3)
- Ubiquitin Transfer: The E2 transfers ubiquitin to the active site cysteine residue within HOIP's RING2 domain, forming a thioester intermediate
- Chain Elongation: The LDD domain positions the acceptor ubiquitin molecule, facilitating nucleophilic attack by the N-terminal methionine amine group on the thioester bond
- Chain Termination: The isopeptide bond formation releases the elongated chain from the catalytic center
This mechanism enables LUBAC to processively build M1-linked polyubiquitin chains on substrate proteins, with HOIL-1L and SHARPIN enhancing the processivity and stability of the complex [4]. Structural analyses reveal that the LDD domain of HOIP provides geometric constraint that specifically favors N-terminal methionine linkage over lysine linkages, accounting for the unique linkage specificity of LUBAC [4].
Table 1: Core Components of the Linear Ubiquitin Chain Assembly Complex (LUBAC)
| Component | Gene | Molecular Weight | Domain Architecture | Primary Function |
|---|---|---|---|---|
| HOIP | RNF31 | 58 kDa | RING1-IBR-RING2-LDD, PUB, UBA, NZF1 | Catalytic subunit with linear chain specificity |
| HOIL-1L | RBCK1 | 123 kDa | RBR, UBL, UBA | Regulatory subunit, stabilizes complex |
| SHARPIN | SHARPIN | 40 kDa | PH, UBL, UBA | Regulatory subunit, enhances E3 activity |
Biological Functions of LUBAC-Generated M1 Ubiquitin
LUBAC-mediated linear ubiquitination regulates multiple key signaling pathways in innate immunity and inflammation:
- NF-κB Signaling: M1-ubiquitin chains conjugated to NEMO/IKKγ and RIPK1 facilitate IKK complex activation by creating scaffolds for TAB/TAK1 complex recruitment and activation [14] [15]
- NOD2 Signaling: Linear ubiquitination of RIPK2 following NOD2 stimulation enhances pro-inflammatory signaling and cytokine production [14]
- TNFR1 Signaling: M1-ubiquitination of complex I components regulates NF-κB activation and cell death decisions [15] [11]
- Cell Death Regulation: LUBAC-mediated linear ubiquitination prevents TNF-induced apoptosis and necroptosis by modifying key components of cell death complexes [15]
Genetic studies in mice demonstrate the critical physiological importance of LUBAC activity. Mutations in Sharpin cause chronic proliferative dermatitis in "cpdm" mice, while HOIP or HOIL-1L deficiency results in embryonic lethality due to aberrant TNFR1-mediated endothelial cell death [4] [11]. In humans, hypomorphic LUBAC mutations cause severe multiorgan autoinflammatory disease, underscoring the essential role of linear ubiquitination in immune homeostasis [4].
Deubiquitination of M1-Linked Chains by OTULIN and CYLD
OTULIN: Specificity and Mechanism
OTULIN (OTU Deubiquitinase With Linear Linkage Specificity; FAM105B) is a 352-amino acid deubiquitinase that exhibits exclusive specificity for Met1-linked polyubiquitin chains, with no detectable activity against other ubiquitin linkage types including the structurally similar K63-linked chains [13]. This remarkable specificity originates from a highly conserved catalytic OTU domain that employs a Cys129/His339/Asn341 catalytic triad and features a specialized ubiquitin-binding pocket that discriminates between different ubiquitin linkages [13].
The molecular mechanism of OTULIN involves substrate-assisted catalysis:
- Autoinhibition: In the absence of ubiquitin, OTULIN's catalytic triad adopts an inactive conformation with Asp336 restricting His339 from its catalytic position
- Ubiquitin Binding: Met1-linked proximal ubiquitin binds to OTULIN's S1' site, orienting the Met1 residue toward the catalytic center
- Activation: Glu16 of the proximal ubiquitin inserts into OTULIN's catalytic center, displacing the inhibitory Asp336 and activating the catalytic His339
- Catalysis: Activated Cys129 performs nucleophilic attack on the isopeptide bond, cleaving the M1 linkage
This Ub-dependent activation mechanism ensures that OTULIN only becomes catalytically active upon engaging its correct substrate, providing an additional layer of specificity beyond mere binding affinity [13]. Mutation of ubiquitin Glu16 to Ala decreases OTULIN's turnover number (kcat) by 240-fold, confirming the critical role of this residue in catalysis [13].
OTULIN contains several functional domains beyond its catalytic OTU domain, including an N-terminal PUB-interacting motif (PIM) that mediates binding to HOIP's PUB domain, and a C-terminal PDZ domain-binding motif that facilitates interactions with endosomal proteins like SNX27 [13] [11]. Phosphorylation of Tyr56 within the PIM domain negatively regulates the OTULIN-HOIP interaction, providing a mechanism for dynamic regulation of OTULIN's access to LUBAC [4].
CYLD: Structure and Dual Specificity
The CYLD lysine 63 deubiquitinase is a ubiquitin-specific protease (USP) family DUB that demonstrates dual specificity for both K63- and M1-linked polyubiquitin chains, with additional activity against K11- and K48-linked chains in vitro [16] [15]. Full-length CYLD comprises 956 amino acids organized into multiple functional domains:
- Three N-terminal CAP-Gly domains that mediate microtubule binding and interaction with the IKK adaptor protein NEMO
- Two proline-rich motifs that potentially interact with SH3 domain-containing proteins
- A TRAF2-binding motif (PVQES) that facilitates substrate recruitment
- A C-terminal USP domain that contains the catalytic center for deubiquitination
Structural studies of the CYLD USP domain reveal how dual specificity for K63- and M1-linked ubiquitin is achieved through a versatile ubiquitin-binding platform that can accommodate different ubiquitin conformations [16] [15]. Unlike OTULIN, CYLD does not rely on substrate-assisted catalysis and shows similar catalytic efficiency against both K63- and M1-linked chains [15].
CYLD activity is regulated through multiple mechanisms, including phosphorylation at S418 by IKK family members, which can either inhibit or activate CYLD depending on cellular context [16]. Additionally, SUMOylation at K40 reduces CYLD-mediated deubiquitination of TRAF2 and TRAF6, while oxidation inhibits catalytic activity, potentially linking CYLD function to cellular redox state [16].
Table 2: Comparison of M1-Linked Ubiquitin Chain Deubiquitinases
| Feature | OTULIN | CYLD |
|---|---|---|
| Gene Name | FAM105B | CYLD |
| Protein Family | OTU | USP |
| Chain Specificity | Exclusive for M1-linked | M1 and K63 (primary), K11, K48 |
| Catalytic Triad | C129, H339, N341 | C601, H871, D889 |
| Mechanism | Substrate-assisted catalysis | Conventional cysteine protease |
| LUBAC Interaction | Direct via HOIP-PUB domain | Indirect via SPATA2 adaptor |
| Non-DUB Functions | Endosomal trafficking (Ub-independent) | Mitotic regulation (DUB-independent) |
| Human Disease | ORAS (Otulipenia) | CYLD Cutaneous Syndrome |
Regulatory Networks and Cross-Talk
OTULIN and CYLD operate within a complex regulatory network that ensures precise control of M1-linked ubiquitination in space and time. Both DUBs interact with LUBAC through distinct mechanisms: OTULIN binds directly to HOIP's PUB domain via its PIM motif, while CYLD is recruited to LUBAC through the adaptor protein SPATA2, which bridges the interaction between CYLD and HOIP [17].
Recent evidence indicates functional cross-talk between these regulatory systems. SPATA2 competes with OTULIN for binding to HOIP, thereby promoting LUBAC autoubiquitination and restricting OTULIN-dependent deubiquitination [17]. This competition creates a dynamic equilibrium where the relative abundance and localization of OTULIN, CYLD, and SPATA2 determine the net level of M1-ubiquitination at signaling complexes.
Genetic studies demonstrate both redundant and unique functions for these DUBs. While CYLD and OTULIN have overlapping substrates in NF-κB signaling, mice deficient in each DUB show distinct phenotypes. OTULIN deficiency causes embryonic lethality due to vascular defects, while CYLD-deficient mice are viable but develop age-dependent conditions [15] [11]. Combined deficiency of CYLD and SPATA2 results in perinatal lethality with enhanced inflammatory signaling, indicating that SPATA2 has OTULIN-dependent functions beyond CYLD recruitment [17].
Experimental Analysis of M1-Linked Ubiquitination
Key Methodologies and Approaches
The study of M1-linked ubiquitination employs specialized methodologies that enable specific detection and quantification of linear ubiquitin chains:
1. Affinity Purification with Linear Ubiquitin-Specific Tools
- Tandem Ubiquitin Binding Entities (TUBEs): Recombinant proteins containing multiple ubiquitin-binding domains that protect ubiquitin chains from DUBs during extraction
- Linear Ubiquitin-Specific Affinity Reagents: Monoclonal antibodies (e.g., HUAB3) or binding domains (e.g., NEMO CC2-ZF) that specifically recognize M1-linked ubiquitin chains
- Protocol: Cells are lysed in denaturing buffer (e.g., 1% SDS) to preserve ubiquitination status, followed by dilution and immunoprecipitation with linear ubiquitin-specific reagents. Bound proteins are analyzed by Western blotting or mass spectrometry.
2. SILAC-Based Quantitative Proteomics for M1-Ubiquitome Analysis
- Principle: Stable Isotope Labeling with Amino acids in Cell culture (SILAC) enables quantitative comparison of protein abundance between experimental conditions
- Workflow:
- Metabolic labeling of cells with light (L-lys0/arg0), medium (L-lys4/arg6), or heavy (L-lys8/arg10) isotopes
- Stimulation under different conditions (e.g., NOD2 ligand treatment)
- Affinity purification of M1-ubiquitinated proteins using linear ubiquitin-specific reagents
- LC-MS/MS analysis and quantitative comparison of enriched proteins
- Application: Identification of RIPK2 as the predominant NOD2-regulated substrate for linear ubiquitination [14]
3. Genetic Manipulation of DUB Activity
- CRISPR/Cas9 Knockout: Generation of CYLD- or OTULIN-deficient cell lines using sgRNAs targeting exon regions
- Catalytic Mutants: Overexpression of catalytically inactive mutants (OTULIN-C129A, CYLD-C601A) to assess DUB-dependent effects
- RNAi-Mediated Depletion: Transient knockdown using siRNA oligonucleotides targeting CYLD or OTULIN coding sequences
Figure 1: Experimental Workflow for M1-Linked Ubiquitination Analysis. This diagram outlines the key methodological steps for studying linear ubiquitination, from sample preparation through functional validation.
Research Reagent Solutions
Table 3: Essential Research Reagents for Studying M1-Linked Ubiquitination
| Reagent Category | Specific Examples | Key Applications | Technical Notes |
|---|---|---|---|
| Linear Ubiquitin-Specific Antibodies | Anti-linear ubiquitin (HUAB3, Millipore), TNF-STAMP M1-specific reagents | Immunofluorescence, Western blotting, Immunoprecipitation | Validate specificity using LUBAC-deficient cells |
| LUBAC Modulators | HOIPIN-1 (HOIP inhibitor), LUBAC expression constructs, siRNA against HOIP/HOIL-1/SHARPIN | Functional studies of linear ubiquitination, Rescue experiments | Titrate inhibitor concentrations carefully |
| DUB Tools | OTULIN-WT and C129A mutants, CYLD-WT and C601A mutants, Recombinant OTULIN/CYLD proteins | In vitro deubiquitination assays, Cellular complementation studies | Use catalytic mutants as negative controls |
| Ubiquitin Variants | Ub(G76S) non-cleavable mutant, Tandem ubiquitin constructs (Ub2-4), Linkage-specific ubiquitin mutants | Biochemical assays, Structural studies, FRET-based assays | G76S prevents cleavage by most DUBs |
| Cell Lines | U2OS/NOD2 reporter cells, OTULIN-/- MEFs, CYLD-/- MEFs, LUBAC-deficient cells | Signaling studies, Genetic screens, Functional validation | Verify genotype regularly |
| Activity Probes | Linear diubiquitin-based activity-based probes (ABPs), Ub-VS derivatives | DUB activity profiling, Inhibitor screening, Cellular imaging | Include controls for specificity |
Pathophysiological Implications and Therapeutic Targeting
Human Disease Associations
Dysregulation of M1-linked ubiquitination machinery underlies several human inflammatory and autoimmune conditions:
OTULIN-Related Autoinflammatory Syndrome (ORAS/ Otulipenia)
- Genetic Basis: Biallelic hypomorphic mutations in OTULIN that reduce but do not completely abolish protein expression
- Clinical Features: Neonatal-onset fever, panniculitis, diarrhea, arthritis, and failure to thrive
- Mechanism: Impaired disassembly of M1-ubiquitin chains leads to excessive TNF-driven NF-κB and ERK signaling, promoting uncontrolled inflammation [11]
- Treatment: TNF inhibitors (e.g., etanercept) show efficacy in managing symptoms
CYLD Cutaneous Syndrome (CCS)
- Genetic Basis: Inherited heterozygous mutations in CYLD that cause truncation of the C-terminal catalytic domain
- Clinical Features: Development of multiple benign skin tumors (cylindromas, trichoepitheliomas, spiradenomas)
- Mechanism: Loss of CYLD-mediated deubiquitination enhances NF-κB and JNK signaling, promoting tumor growth in hair follicle and skin appendages [16]
Neurodegenerative Disorders
- Association: Specific gain-of-function CYLD mutations linked to frontotemporal dementia and amyotrophic lateral sclerosis
- Mechanism: Hyperactive deubiquitination disrupts protein quality control and promotes TDP-43 aggregation [16]
Additionally, somatic mutations in LUBAC components and regulatory DUBs are increasingly recognized in various cancers, highlighting the importance of balanced linear ubiquitination for cellular homeostasis [4] [11].
Therapeutic Targeting Strategies
Several strategic approaches are being explored to target the linear ubiquitination pathway for therapeutic benefit:
1. LUBAC Inhibition
- Small Molecule Inhibitors: HOIPIN compounds that target the LDD domain of HOIP and inhibit linear chain formation
- Therapeutic Application: Potential for treating autoimmune conditions characterized by excessive NF-κB activation
- Challenges: Achieving specificity over other E3 ligases and minimizing toxicity
2. DUB-Targeted Therapies
- OTULIN Activation: Strategies to enhance OTULIN activity or expression could dampen pathological M1-ubiquitination in autoinflammatory diseases
- CYLD Inhibition: In specific contexts where enhanced CYLD activity contributes to disease pathogenesis
- Considerations: Tissue-specific delivery and temporal control of modulation
3. Hybrid Chain Targeting
- Approach: Development of reagents that specifically target M1/K63 hybrid ubiquitin chains that are particularly effective at activating NF-κB signaling
- Rationale: These hybrid chains are naturally occurring and show enhanced signaling capacity in inflammatory pathways
Figure 2: Regulatory Network of M1-Linked Ubiquitination in Immune Signaling. This diagram illustrates the core pathway from receptor stimulation to NF-κB activation, highlighting points of regulation by OTULIN and CYLD deubiquitination activities.
The precise mechanisms governing M1-linked ubiquitin chain assembly by LUBAC and disassembly by OTULIN and CYLD represent a crucial regulatory axis in inflammatory signaling and cell fate determination. The unique specificity of OTULIN for linear chains and the dual specificity of CYLD create complementary regulatory layers that ensure appropriate signal duration and magnitude. Structural insights into the molecular mechanisms of these enzymes—particularly OTULIN's substrate-assisted catalysis—provide foundations for rational drug design.
Key challenges and future directions in this field include:
- Understanding Context-Specific Regulation: Determining how OTULIN and CYLD achieve substrate specificity in different signaling contexts and cell types
- Elucidating Ubiquitin-Independent Functions: Characterizing the emerging non-catalytic roles of OTULIN in endosomal trafficking and other cellular processes
- Developing Clinically Useful Modulators: Advancing specific small molecule regulators of LUBAC, OTULIN, and CYLD for therapeutic applications
- Exploring Tissue-Specific Functions: Using tissue-specific knockout models to understand how linear ubiquitination regulates homeostasis in different organs
The rapidly expanding knowledge of M1-linked ubiquitination continues to reveal unexpected complexity in this essential signaling system. As research methodologies advance—particularly in structural biology, proteomics, and genetic manipulation—our understanding of how linear ubiquitination controls immune signaling and cellular fate will undoubtedly deepen, opening new therapeutic avenues for inflammatory disease, cancer, and immune disorders.
The activation of the nuclear factor-κB (NF-κB) pathway is a cornerstone of innate and adaptive immunity, orchestrating inflammatory responses to pathogens and cellular stress. This whitepaper delineates the critical regulatory function of M1-linked linear ubiquitination in fine-tuning NF-κB signaling downstream of Toll-like Receptors (TLRs) and the Tumor Necrosis Factor Receptor (TNFR). As a unique post-translational modification, M1-ubiquitin chains are exclusively assembled by the Linear Ubiquitin Chain Assembly Complex (LUBAC) [18] [19]. We detail the mechanisms by which LUBAC-generated linear ubiquitin chains act as pivotal molecular scaffolds that facilitate the recruitment and activation of key kinase complexes, ultimately ensuring robust NF-κB–driven gene transcription. Furthermore, this guide integrates quantitative biochemical data, provides detailed experimental methodologies for studying these pathways, and visualizes the complex protein interactions through standardized diagrams. Given its central role in inflammation, the LUBAC-M1-ubiquitin axis presents a compelling target for therapeutic intervention in autoimmune, inflammatory, and oncological diseases.
Ubiquitination is a versatile post-translational modification where a 76–amino acid polypeptide, ubiquitin, is covalently attached to substrate proteins. Unlike other polyubiquitin linkages, M1-linked (or linear) ubiquitin chains are formed through a peptide bond between the C-terminal glycine of one ubiquitin and the N-terminal methionine (M1) of the next [18]. The discovery that the multi-protein complex LUBAC is the sole known cellular E3 ligase capable of generating M1-linked chains placed this modification at the center of inflammatory signaling research [19]. LUBAC comprises three core subunits: HOIP (the catalytic subunit), HOIL-1L, and SHARPIN [18] [19].
In the context of immune signaling, NF-κB is a master regulator of inflammation, controlling the expression of cytokines, chemokines, and anti-apoptotic genes. The canonical NF-κB activation pathway, triggered by ligands for TNFR1 and TLRs, converges on the activation of the IκB kinase (IKK) complex. This complex, consisting of the catalytic subunits IKKα and IKKβ and the regulatory subunit NEMO (IKKγ), phosphorylates the inhibitory protein IκBα, targeting it for degradation and thereby releasing the NF-κB dimer (typically p50/RelA) for nuclear translocation [20] [21]. This whitpaper will dissect how M1-ubiquitination, strategically deployed by LUBAC, serves as an essential molecular platform that integrates and amplifies signals from diverse receptors to precisely control this critical pathway.
Molecular Mechanisms of M1-Ubiquitination in NF-κB Signaling
M1-Ubiquitination in TNFR1 Signaling
Upon binding of TNF-α to TNFR1, the receptor trimerizes and initiates the assembly of a primary signaling complex (Complex I). The death domain (DD) of TNFR1 recruits the adaptor protein TRADD, which in turn recruits TRAF2 and RIPK1 [22]. It is at this complex that LUBAC is recruited, where it subsequently conjugates M1-linked ubiquitin chains to key components, including RIPK1 and NEMO [19].
- Scaffold for Kinase Activation: The M1-ubiquitin chains deposited by LUBAC act as a docking platform for the TAK1 (TGF-β-activated kinase 1) complex and the IKK complex. Both complexes contain ubiquitin-binding domains that specifically recognize linear ubiquitin chains. This recruitment brings TAK1 into close proximity with the IKK complex, enabling TAK1-mediated phosphorylation and activation of IKKβ [21].
- Enhancement of Signaling Fidelity: The presence of M1-ubiquitin chains stabilizes Complex I and promotes full IKK activation. This step is critical for the subsequent phosphorylation and proteasomal degradation of IκBα, which unleashes NF-κB to enter the nucleus and activate pro-survival and inflammatory genes [22] [21].
The diagram below illustrates the pivotal role of LUBAC and M1-ubiquitin in the TNFR1 signaling pathway.
M1-Ubiquitination in TLR Signaling
TLR signaling, particularly through the adapter MyD88, shares a common requirement for M1-ubiquitin with the TNFR1 pathway. Upon TLR engagement (e.g., by LPS), the signal propagates through MyD88, IRAK kinases, and the E3 ligase TRAF6 [21]. TRAF6 catalyzes the formation of K63-linked ubiquitin chains, which in turn recruit the TAK1 and IKK complexes.
- Synergy with K63 Ubiquitination: LUBAC is recruited to the activated TLR complex and generates M1-linked ubiquitin chains alongside K63 chains. The M1 chains provide a high-affinity binding site for NEMO, whose UBAN domain has a specific affinity for linear ubiquitin. This synergistic action ensures robust and specific activation of the IKK complex [18] [21].
- Role in Sterile Inflammation: Beyond pathogen infection, M1-ubiquitination is a conserved response to sterile stressors like hypoxia, oxidative stress, and mechanical stress. Studies in Drosophila melanogaster and human cells show that the LUBEL/LUBAC-dependent formation of M1-Ub chains is essential for NF-κB activation and cell survival under these stress conditions, linking this pathway directly to inflammation in chronic diseases and cancer [18].
The following diagram outlines the integration of M1-ubiquitination into the TLR signaling cascade.
Quantitative Data in M1-Ubiquitin Research
The study of M1-ubiquitination relies on quantitative data to elucidate binding affinities, kinetic parameters, and functional outcomes. The tables below summarize key quantitative findings and the regulatory landscape of this pathway.
Table 1: Key Biophysical and Functional Data in M1-Ubiquitin Signaling
| Protein/Complex | Key Interacting Partner | Affinity (Kd) / Method | Functional Outcome | Experimental Context | Source |
|---|---|---|---|---|---|
| NEMO (UBAN domain) | M1-linked diUb | ~1-10 µM (SPR) | Recruitment of IKK complex to M1-ubiquitinated scaffolds | Recombinant proteins | [19] |
| LUBAC (HOIP) | Ubiquitin (acceptor) | N/A (E3 Catalysis) | Catalytic formation of M1-Ub chains | In vitro ubiquitination assay | [18] [19] |
| M1-Ub Chain Formation | Hypoxia/Oxidative Stress | >2-fold induction (M1-TUBE pulldown) | Activation of NF-κB; Protection from stress-induced cell death | Drosophila and human Caco2 cells | [18] |
| Immune Surface Interactions | Various receptor-ligand pairs | Center: Low µM range (SPR) | Transition to higher-affinity interactions in inflamed state | Systematic human interactome screen | [23] |
Table 2: Regulatory Enzymes and Genetic Links to Disease
| Component | Type | Function | Associated Human Diseases | Source |
|---|---|---|---|---|
| HOIP (RBR-LDD domain) | E3 Ubiquitin Ligase | Catalytic core of LUBAC; forms M1-Ub chains | Autoimmunity, Immunodeficiencies | [18] [19] |
| OTULIN (Gumby) | Deubiquitinase (DUB) | Hydrolyzes M1-Ub chains; negative regulator | Autoinflammatory syndrome (OTULINopathy) | [19] |
| A20 (TNFAIP3) | Deubiquitinase & E3 Ligase | Negative regulator; edits K63 and M1 chains | B-cell lymphomas, Autoimmune disorders | [20] [19] |
| CYLD | Deubiquitinase (DUB) | Cleaves K63 and M1-linked Ub chains; negative regulator | Cylindromatosis (skin tumors) | [19] |
Experimental Protocols for Investigating M1-Ubiquitination
To empirically investigate the role of M1-ubiquitination in NF-κB signaling, the following methodologies are essential.
Detecting M1-Ubiquitin Chains in Cellular Signaling
Purpose: To detect and quantify the inducible formation of endogenous M1-ubiquitin chains in cells upon pathway stimulation (e.g., with TNF-α or LPS).
Reagents:
- M1-Specific Tandem Ubiquitin-Binding Entity (M1-TUBE): A recombinant protein tool with high avidity and specificity for M1-linked ubiquitin chains, used for immunoaffinity purification [18].
- Cell Lysis Buffer: Containing protease inhibitors, deubiquitinase (DUB) inhibitors (e.g., N-Ethylmaleimide), and SDS to preserve labile ubiquitin modifications.
- Stimuli: Recombinant human TNF-α (e.g., 10-50 ng/mL) or ultrapure LPS (e.g., 100 ng/mL).
- Antibodies: For western blotting: anti-UB (linear) specific antibody (e.g., from Millipore), anti-RIPK1, anti-NEMO, anti-IκBα, and anti-phospho-IκBα.
Procedure:
- Stimulation and Lysis: Culture mammalian cells (e.g., HEK293, HeLa, or primary macrophages) and stimulate with TNF-α or LPS for a time-course (e.g., 0, 5, 15, 30 min). Immediately lyse cells in pre-heated SDS-containing buffer and boil samples to inactivate DUBs.
- M1-Ub Chain Enrichment: Dilute lysates to reduce SDS concentration and incubate with GST-tagged M1-TUBE pre-bound to glutathione-sepharose beads for 2-4 hours at 4°C.
- Washing and Elution: Wash beads extensively with mild wash buffer. Elute bound proteins with SDS-PAGE sample buffer.
- Analysis: Analyze eluates by SDS-PAGE and western blotting using the anti-linear ubiquitin antibody to detect enriched M1-chains. Reprobing the blot for signaling components like RIPK1 or NEMO can identify proteins modified with or co-purifying with M1-chains.
Assessing NF-κB Activation Status
Purpose: To monitor the functional consequence of M1-ubiquitination by measuring NF-κB pathway activation at multiple levels.
Methods:
- IKK Kinase Assay: Immunoprecipitate the IKK complex from cell lysates and perform an in vitro kinase assay using recombinant IκBα or GST-IκBα as a substrate. Detection of phosphorylated IκBα by western blot indicates IKK activity [21].
- IκBα Degradation and Phosphorylation Immunoblot: A standard and straightforward assay. Whole-cell lysates from stimulated cells are immunoblotted for total IκBα and phospho-IκBα (Ser32/36). Rapid degradation and transient phosphorylation of IκBα are hallmarks of canonical NF-κB activation [20] [21].
- Nuclear Translocation and DNA Binding:
- Immunofluorescence (IF): Fix and permeabilize stimulated cells, then stain with an antibody against the p65 (RelA) subunit of NF-κB. Monitor translocation from cytoplasm to nucleus using fluorescence microscopy. This is ideal for single-cell analysis and capturing heterogeneity [20].
- Electrophoretic Mobility Shift Assay (EMSA): Prepare nuclear extracts and incubate with a radiolabeled DNA probe containing a κB consensus sequence. A gel shift indicates NF-κB DNA-binding activity. While a classic technique, it has been largely superseded by methods more amenable to quantification.
- Target Gene Expression Analysis: Extract total RNA and analyze mRNA levels of classic NF-κB target genes (e.g., IL6, IL8, TNFAIP3/A20) using quantitative RT-PCR (qPCR). This measures the ultimate transcriptional output of the pathway [20].
The workflow for a comprehensive NF-κB activation analysis is depicted below.
The Scientist's Toolkit: Key Research Reagents and Models
Table 3: Essential Research Tools for M1-Ubiquitin and NF-κB Studies
| Reagent / Model | Type | Key Function / Application | Key Feature |
|---|---|---|---|
| M1-TUBE | Recombinant Protein | Affinity purification of M1-linked ubiquitin chains from cell lysates. | High specificity over other ubiquitin chain types. |
| Anti-Linear Ubiquitin Antibody | Antibody | Detection of M1-ubiquitin chains by western blot, IF, and IHC. | Distinguishes M1-linkage from K-linked chains. |
| LUBAC Inhibitors | Small Molecule | Pharmacological inhibition of HOIP catalytic activity (RBR domain). | Tool for acute disruption of M1-ubiquitination. |
| OTULIN Wild-type & Mutant | Recombinant Protein / cDNA | Active (WT) and catalytically inactive (Mutant) DUB to manipulate cellular M1-Ub levels. | Critical for gain/loss-of-function studies. |
| Drosophila melanogaster LUBEL mutant | Genetic Model | In vivo study of M1-ubiquitination in development, immunity, and stress responses. | HOIP homolog; conserved pathway. |
| SHARPIN-deficient mice (cpdm) | Genetic Model | In vivo model of chronic LUBAC dysfunction to study systemic inflammation. | Spontaneous dermatitis and immunodeficiency. |
The discovery of LUBAC and M1-linked ubiquitination has unveiled a critical regulatory layer in NF-κB activation that is fundamental to inflammatory signaling via TNFR and TLR pathways. This whitepaper has detailed the molecular mechanics, quantitative parameters, and experimental approaches that define this field. The precise manipulation of this pathway—through inhibition of HOIP or modulation of regulatory DUBs like OTULIN and A20—holds immense therapeutic potential. Future research will undoubtedly focus on developing highly specific drugs targeting LUBAC, understanding the crosstalk between M1-ubiquitination and other post-translational modifications (such as the recently discovered MARUbylation [24]), and elucidating the role of linear ubiquitination in the context of the broader immune interactome [23]. Integrating these insights will be crucial for devising novel treatment strategies for a wide spectrum of inflammatory and autoimmune pathologies.
M1-linked linear ubiquitin chains, once primarily studied in the context of pathogen infection, are now recognized as critical signaling hubs in sterile inflammation. This whitepaper synthesizes current research demonstrating how hypoxia and oxidative stress activate the M1-ubiquitination machinery, particularly through mitochondrial reactive oxygen species (mitoROS), to drive NF-κB-mediated inflammatory signaling. We detail the molecular mechanisms, experimental methodologies, and key research tools essential for investigating this pathway, providing a technical foundation for researchers exploring therapeutic interventions in inflammatory diseases and cancer.
Ubiquitination is a sophisticated post-translational modification system that regulates protein stability, activity, and interactions through covalent attachment of ubiquitin molecules. Among the various ubiquitin chain linkages, M1-linked linear ubiquitin chains (where the C-terminal glycine of one ubiquitin connects to the N-terminal methionine of another) have emerged as specialized signaling scaffolds that potently activate inflammatory pathways [25]. Initially characterized in pathogen-responsive signaling complexes such as the linear ubiquitin chain assembly complex (LUBAC) in mammals [26], M1-ubiquitination is now recognized as a crucial mechanism in sterile inflammation – inflammatory responses occurring in the absence of pathogens.
The NF-κB pathway serves as a primary signaling hub regulated by M1-ubiquitination, with chains facilitating critical protein-protein interactions that lead to IKK complex activation and subsequent pro-inflammatory gene expression [27] [25]. Beyond infection contexts, recent research has illuminated how cellular stress conditions – particularly hypoxia and oxidative stress – exploit this molecular machinery to initiate and sustain inflammation in conditions ranging from chronic inflammatory diseases to cancer [28] [29].
Molecular Mechanisms of M1-Ubiquitination in Sterile Stress
Hypoxia-Induced M1-Ubiquitination via Mitochondrial ROS
Hypoxia represents a fundamental cellular stress that occurs in pathological contexts including cancer, ischemic conditions, and chronic inflammatory diseases. Recent preprint findings reveal that hypoxia-induced mitoROS triggers significant accumulation of M1-ubiquitin chains, which subsequently activates NF-κB signaling and expression of its target genes [28]. This mechanism connects mitochondrial dysfunction to inflammatory signaling through ubiquitination, representing a paradigm shift in understanding how metabolic stress translates to inflammation.
Table 1: Key Findings on Hypoxia-Induced M1-Ubiquitination
| Experimental Finding | Significance | Reference |
|---|---|---|
| Hypoxia induces M1-ubiquitin chain accumulation | Links mitochondrial stress to inflammatory signaling | [28] |
| Mitochondrial ROS necessary for M1-chain formation | Identifies mitoROS as trigger mechanism | [28] |
| M1-chains activate NF-κB signaling | Connects hypoxia to inflammation transcription program | [28] |
| Hypoxia-induced mitophagy occurs independently of ubiquitination | Suggests specialized role for M1-ubiquitination in signaling | [28] |
Conservation Across Stressors and Species
The role of M1-ubiquitination in sterile inflammation extends beyond hypoxia and is evolutionarily conserved. Research in Drosophila melanogaster demonstrates that the LUBEL enzyme (linear ubiquitin E3 ligase, the HOIP orthologue) catalyzes M1-ubiquitin chain formation in response to diverse stressors including hypoxia, oxidative stress (paraquat-induced), and mechanical stress [29]. This pathway activates the Drosophila Imd pathway, leading to NF-κB (Relish) activation and stress adaptation. The conservation of this mechanism from flies to mammals (where LUBAC performs analogous functions) underscores the fundamental importance of M1-ubiquitination in stress responses [25] [29].
The molecular circuitry involves a cascade where stress signals lead to LUBEL/LUBAC-mediated M1-ubiquitination of IKKγ (Kenny in flies, NEMO in mammals), facilitating kinase complex activation and NF-κB nuclear translocation [25] [29]. This pathway operates independently of pathogen-sensing receptors, instead responding directly to metabolic and cellular disturbances.
Experimental Evidence and Quantitative Data
Genetic Evidence from Model Systems
Studies in Drosophila provide compelling genetic evidence for the essential role of M1-ubiquitination in stress survival. LUBEL-deficient flies exhibit significantly reduced survival under both hypoxic conditions and paraquat-induced oxidative stress compared to wild-type controls [29]. This phenotypic evidence strongly supports the physiological importance of the pathway in stress adaptation.
The genetic hierarchy within this pathway has been delineated through systematic analysis, revealing that LUBEL functions downstream of stress sensing but upstream of the IKK complex, requiring intact Tak1, Diap2, Kenny (IKKγ), and Dredd signaling components for sterile inflammatory responses [29].
Table 2: Survival Analysis in Stress Models with M1-Ubiquitination Deficiencies
| Stress Condition | Experimental Model | Key Finding | Molecular Dependence |
|---|---|---|---|
| Hypoxia | Drosophila LUBEL mutants | Reduced survival | Imd pathway components |
| Oxidative stress (paraquat) | Drosophila LUBEL mutants | Reduced survival | Tak1, Diap2, Kenny, Dredd |
| Oral bacterial infection | Drosophila LUBEL mutants | Reduced survival & pathogen clearance | IMD pathway, DIAP2 |
| Systemic infection | Drosophila LUBEL mutants | Normal survival | Pathway not required |
Hybrid Ubiquitin Chains in Innate Immune Signaling
Beyond pure M1-linked chains, recent research has revealed the prevalence and significance of Lys63/Met1-hybrid ubiquitin chains in innate immune signaling. These hybrid chains, containing both K63-linked and M1-linked ubiquitin oligomers, are rapidly formed when multiple innate immune signaling networks are activated, including TNFR1/TRADD, TLR3/TRIF, and NOD1/RIP2 pathways [26].
The formation of these hybrid chains on key signaling adaptors such as RIP1 (TNFR1 pathway) and RIP2 (NOD1 pathway) suggests they may provide synergistic signaling advantages over homogeneous chains, potentially creating more diverse platforms for recruiting proteins with different ubiquitin-binding domains [26].
Experimental Protocols and Methodologies
Detecting M1-Ubiquitin Chains in Stress Models
The following methodology has been successfully employed to detect and quantify M1-ubiquitin chain formation in response to sterile stressors:
Protocol: M1-Ubiquitin Chain Pull-Down Assay
- Cell Lysis and Preparation: Lyse cells or tissue samples in modified RIPA buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS) supplemented with 10 mM N-ethylmaleimide (NEM) to preserve ubiquitin chains, complete protease inhibitors, and 1 mM PMSF.
- Ubiquitin Chain Enrichment: Incubate cleared lysates with GST-tagged ubiquitin-binding domain (UBD) probes specifically recognizing M1-linkages:
- Washing and Elution: Wash beads extensively with lysis buffer, then elute bound proteins with SDS sample buffer.
- Specificity Controls:
- Treat samples with recombinant OTULIN (M1-specific deubiquitinase) to verify chain linkage
- Compare to vOTU treatment (cleaves all chain types except M1-linked)
- Use lysates from LUBEL/LUBAC-deficient cells as negative controls
- Detection: Analyze by SDS-PAGE and western blotting using anti-ubiquitin antibodies capable of recognizing linear epitopes.
Stress Induction Conditions:
- Hypoxia: Place cells in hypoxia chamber (1% O₂, 5% CO₂, balance N₂) for 2-8 hours
- Oxidative Stress: Treat with paraquat (100-500 µM) or hydrogen peroxide (100-400 µM) for 1-4 hours
- Mechanical Stress: Apply cyclic stretch (10-15% elongation) using flexible membrane systems
Functional Validation of M1-Ubiquitination in NF-κB Signaling
To establish the functional consequences of stress-induced M1-ubiquitination:
Protocol: NF-κB Activation Assay
- Genetic Manipulation:
- Knockdown or knockout LUBEL/LUBAC components (HOIP, HOIL-1, SHARPIN) using RNAi or CRISPR-Cas9
- Overexpress catalytic domains of LUBEL to examine gain-of-function effects
- Reporter Assay: Transfect NF-κB luciferase reporter constructs along with Renilla control for normalization
- Stress Application: Expose cells to hypoxia or oxidative stress as described above
- Readout:
- Measure luciferase activity to quantify NF-κB transcriptional activation
- Analyze nuclear translocation of NF-κB subunits (p65/RelA) by immunofluorescence or subcellular fractionation
- Quantify expression of endogenous NF-κB target genes (IL-8, IL-6, TNF-α) by RT-qPCR
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Studying M1-Ubiquitination in Sterile Inflammation
| Reagent/Tool | Specific Example | Function/Application | Experimental Use |
|---|---|---|---|
| M1-chain Binders | GST-NEMO-UBAN fusion protein | High-affinity M1-ubiquitin chain recognition | Pull-down assays, chain detection |
| Linkage-specific DUBs | Recombinant OTULIN | M1-chain hydrolysis (specificity control) | Verification of M1-linkages in samples |
| E3 Ligase Tools | LUBEL/HOIP expression constructs | M1-ubiquitin chain formation | Gain-of-function studies |
| E3 Ligase Inhibitors | HOIP inhibitors (e.g., HBX) | Block M1-chain assembly | Loss-of-function studies |
| Genetic Models | LUBEL-deficient Drosophila | In vivo functional analysis | Stress survival assays, genetic screening |
| Detection Antibodies | Linear ubiquitin-specific antibodies | M1-chain immunodetection | Western blot, immunofluorescence |
| Stress Inducers | Paraquat, Hypoxia chambers | Induce oxidative stress and hypoxia | Pathway activation studies |
| Signaling Reporters | NF-κB luciferase reporters | Measure pathway activation | Functional consequence assessment |
Discussion and Research Implications
The emerging role of M1-ubiquitination in sterile inflammation represents a significant expansion of our understanding of ubiquitin signaling beyond its traditional domains. The mechanistic link between mitochondrial stress, M1-ubiquitin chain formation, and NF-κB activation provides a unified framework for understanding how diverse stressors converge on a common inflammatory pathway [28] [29]. This has profound implications for numerous pathological conditions:
Therapeutic Targeting Opportunities: Components of the M1-ubiquitination pathway represent attractive targets for inflammatory diseases and cancer. The non-redundant functions of LUBAC components and the specificity of M1-chains for inflammatory signaling suggest potential for targeted interventions with reduced side effects compared to broader immunosuppressants.
Cancer Microenvironment: Hypoxia is a hallmark of solid tumors, and the hypoxia-mitoROS-M1-ubiquitination axis may contribute to the chronic inflammatory microenvironment that supports tumor progression and resistance to therapy. Targeting this pathway may enhance current cancer immunotherapies [30].
Chronic Inflammatory Diseases: Conditions characterized by sterile inflammation, including metabolic diseases, neurodegenerative disorders, and autoimmune conditions, may involve dysregulation of stress-induced M1-ubiquitination pathways. Modulating this pathway could potentially resolve inflammation without compromising anti-pathogen immunity.
M1-linked ubiquitination has emerged as a critical signaling mechanism that extends far beyond its initial characterization in pathogen response. In sterile inflammation triggered by hypoxia and oxidative stress, M1-ubiquitin chains serve as essential molecular switches that activate NF-κB signaling and coordinate cellular adaptation to stress. The conservation of this pathway from Drosophila to mammals underscores its fundamental importance in physiology and disease. Continued investigation of the molecular mechanisms governing stress-induced M1-ubiquitination will undoubtedly yield novel insights into inflammatory pathogenesis and reveal new therapeutic opportunities for a range of human diseases.
The cellular response to infection and injury is governed by a precise equilibrium between pro-inflammatory signaling and programmed cell death. Recent advances illuminate M1-linked/linear ubiquitin chains as a central regulatory mechanism coordinating this balance. This whitepaper synthesizes current research demonstrating how the linear ubiquitin chain assembly complex (LUBAC) and specific E3 ubiquitin ligases direct macrophage polarization, inflammatory pathway activation, and cell death decisions through ubiquitin-dependent signaling. Dysregulation of these processes contributes to chronic inflammation, autoimmunity, and sepsis pathophysiology, revealing novel therapeutic targets for immune-related disorders. Technical methodologies for investigating ubiquitin signaling and experimental approaches for targeting these pathways are discussed in depth.
Protein ubiquitination, the post-translational attachment of ubiquitin to substrate proteins, serves as a master regulator of innate immune responses. The M1-linked linear ubiquitin chain, characterized by ubiquitin monomers connected through methionine-1 residues, has emerged as a critical signaling scaffold in inflammatory pathways and cell fate decisions. Unlike degradative K48-linked chains, M1 linkages primarily function as molecular scaffolds that recruit signaling complexes to activate NF-κB, regulate cell death pathways, and direct macrophage polarization [31] [32].
The generation of M1 ubiquitin chains is catalyzed by the linear ubiquitin chain assembly complex (LUBAC), consisting of HOIP, HOIL-1L, and SHARPIN components. HOIP serves as the catalytic core possessing unique determinant residues that specify linear/M1 linkage formation [31]. Recent research has positioned LUBAC-generated M1 chains as pivotal integrators balancing inflammatory signaling against cell death execution, particularly in macrophages where they regulate the transition between pro-inflammatory M1 and anti-inflammatory M2 states [32] [33].
This technical review examines the molecular mechanisms through which M1 ubiquitination regulates the critical balance between inflammatory signaling and cell death, with emphasis on experimental approaches for investigating these pathways and their therapeutic implications for immune disorders.
Molecular Mechanisms of M1 Ubiquitin Signaling
LUBAC and STING-Mediated NF-κB Activation
Recent research has elucidated a novel mechanism whereby stimulator of interferon genes (STING) activation induces HOIP-mediated synthesis of M1 ubiquitin chains to stimulate NF-κB signaling. STING activation by cyclic dinucleotides recruits LUBAC to LC3B-associated Golgi membranes where ubiquitin is localized. This recruitment facilitates M1-linked ubiquitin chain formation independent of STING-induced LC3B lipidation, representing a distinct pathway for innate immune activation [31].
Table 1: Key Proteins in M1 Ubiquitin-Dependent Inflammatory Signaling
| Protein/Complex | Function | Regulatory Role | Pathway Association |
|---|---|---|---|
| LUBAC (HOIP) | Catalytic component of E3 ligase complex | Synthesizes M1-linked ubiquitin chains | STING, NF-κB, TNF signaling |
| STING | Innate immune sensor for cyclic dinucleotides | Recruits LUBAC to Golgi membranes | NF-κB, IRF3 activation |
| OTULIN | Deubiquitinase | Hydrolyzes linear/M1 ubiquitin chains | Prevents spontaneous NF-κB activation |
| A20 (TNFAIP3) | Ubiquitin-editing enzyme | Removes activating chains, adds degradative chains | Negative feedback in NF-κB signaling |
| Praja2 | E3 ubiquitin ligase | Ubiquitylates MFHAS1 without degradation | Promotes M1 polarization via JNK/p38 |
| NEMO (IKBKG) | Regulatory subunit of IKK complex | Binds M1 ubiquitin chains via UBAN domain | NF-κB activation |
The functional outcome of STING-induced M1 ubiquitination is the activation of both NF-κB and IRF3 signaling pathways, coordinating inflammatory and interferon responses. Genetic ablation of HOIP prevents M1 ubiquitin chain formation and significantly reduces both NF-κB and IRF3 signaling in human monocytes and mouse bone marrow-derived macrophages, without affecting STING activation per se [31]. This demonstrates the essential positioning of LUBAC downstream of STING activation but upstream of transcription factor signaling.
Ubiquitination in Macrophage Polarization and Plasticity
Macrophage polarization represents a critical interface between inflammatory signaling and cell fate decisions. Ubiquitination, particularly through M1 chains, serves as a fundamental mechanism directing macrophage functional plasticity:
M1 Polarization: Pro-inflammatory M1 macrophage polarization is regulated by multiple E3 ubiquitin ligases. Praja2 ubiquitylates MFHAS1 without promoting its degradation, instead activating TLR2-mediated JNK/p38 pathways that drive M1 polarization and enhance production of IL-6, TNF-α, and iNOS [34]. The E3 ligase TRIM21 promotes M1 polarization post-myocardial infarction, with TRIM21-deficient mice exhibiting smaller infarcts and increased M2 macrophages [32].
M2 Polarization: Anti-inflammatory M2 polarization involves distinct ubiquitin machinery. Pellino1 enhances STAT3 stability through ubiquitination, supporting IL-10-induced M2 polarization [32]. MARCH1, induced by IL-10, ubiquitinates MHC II and CD86, targeting them for degradation and enforcing an immunosuppressive state [32].
Plasticity Regulation: Deubiquitinases provide critical counter-regulation. CYLD removes K63-linked chains from NF-κB and JNK adaptors, curbing M1 cytokine output, while OTULIN hydrolyzes linear ubiquitin chains on TLR and TNF signaling adaptors [32] [33]. OTULIN deficiency causes ligand-independent NF-κB activation and severe inflammation, mimicking human ORAS (OTULIN-related autoinflammatory syndrome) [32].
Cell Death Pathways in Inflammation and Immunity
Cell death represents the ultimate cell fate decision with profound implications for inflammatory outcomes. The interplay between ubiquitination and cell death pathways creates critical regulatory nodes that determine disease progression, particularly in sepsis and chronic inflammation.
Table 2: Cell Death Pathways in Inflammatory Regulation
| Cell Death Pathway | Key Mediators | Inflammatory Outcome | Role in Sepsis/Inflammation |
|---|---|---|---|
| Apoptosis | Caspase-3, -8, -9; Bcl-2 family | Anti-inflammatory; promotes tolerance | Lymphocyte apoptosis induces immunosuppression; reduced neutrophil apoptosis exacerbates inflammation |
| Pyroptosis | Caspase-1, -4, -5; Gasdermin D | Highly inflammatory; IL-1β/IL-18 release | Elevated in sepsis; contributes to cytokine storm |
| Necroptosis | RIPK1, RIPK3, MLKL | Pro-inflammatory; DAMP release | Widespread necrosis exacerbates sepsis inflammation |
| Ferroptosis | Iron accumulation, lipid peroxidation | Inflammatory; HMGB1, cfDNA release | Emerging role in sepsis pathophysiology |
| NETosis | Neutrophil extracellular traps | Highly inflammatory; antimicrobial | Triggers autoimmunity in SLE, RA |
Apoptosis in Immune Regulation
Apoptosis, particularly of immune cells, plays a dual role in inflammatory regulation. Neutrophil apoptosis followed by efferocytosis promotes resolution of inflammation, as apoptotic cells expose phosphatidylserine (PS) on their surface, serving as "eat-me" signals that drive macrophages toward anti-inflammatory, pro-resolving phenotypes [35]. In sepsis, however, apoptosis becomes dysregulated: neutrophil apoptosis decreases, prolonging inflammation, while lymphocyte apoptosis increases, contributing to immunosuppression [36] [35].
The ubiquitin system regulates apoptotic signaling through multiple mechanisms. A20 terminates NF-κB signaling by removing activating ubiquitin chains from adaptor proteins and adding degradative chains, while Pellino1 ubiquitinates and stabilizes STAT3 to promote anti-inflammatory signaling [32]. These ubiquitin-dependent checks balance the pro- and anti-apoptotic signals that determine immune cell fate during inflammation.
Inflammatory Cell Death in Sepsis and Chronic Inflammation
Pyroptosis, necroptosis, and NETosis represent highly inflammatory cell death modalities that contribute significantly to sepsis pathophysiology and chronic inflammatory diseases:
Pyroptosis proceeds through canonical (caspase-1) and non-canonical (caspase-4/5/11) pathways, culminating in Gasdermin D-mediated pore formation and release of IL-1β, IL-18, and other DAMPs [36]. The deubiquitinase BRCC3 removes K48/K63 ubiquitin from NLRP3, permitting inflammasome assembly and pyroptosis execution [32].
Necroptosis represents a regulated form of necrosis initiated by receptor-mediated interactions, resulting in uncontrolled release of cytosolic DAMPs including HMGB1, extracellular RNAs, and cell-free DNA [35].
NETosis involves the release of web-like chromatin structures containing DNA, histones, and antimicrobial peptides. While microbiocidal, excessive NETosis contributes to tissue damage and triggers autoimmune responses in conditions like systemic lupus erythematosus and rheumatoid arthritis [35].
The ubiquitin code determines cell death modality selection, with K63-linked and M1-linked chains often promoting survival signaling, while specific chain types can facilitate inflammatory death execution.
Experimental and Technical Approaches
Quantitative Ubiquitylomics Methodologies
Advanced mass spectrometry-based techniques enable comprehensive analysis of ubiquitin signaling. Data-independent acquisition (DIA) methods coupled with diGly antibody-based enrichment have revolutionized ubiquitinome profiling:
Key methodological considerations:
- Peptide input: 1mg peptide material using 31.25μg anti-diGly antibody optimal for enrichment [37]
- Fractionation: Basic reversed-phase chromatography into 96 fractions concatenated to 8 pools improves coverage [37]
- K48-peptide handling: Separate processing of fractions containing abundant K48-linked ubiquitin-chain derived diGly peptide reduces competition during enrichment [37]
- DIA parameters: 46 precursor isolation windows with MS2 resolution of 30,000 provides optimal performance for diGly peptide analysis [37]
This optimized workflow identifies approximately 35,000 distinct diGly peptides in single measurements of proteasome inhibitor-treated cells—doubling the coverage achievable with data-dependent acquisition methods [37]. The improved quantitative accuracy (45% of diGly peptides with CVs <20% vs. 15% with DDA) enables robust systems-wide investigations of ubiquitin signaling dynamics [37].
Research Reagent Solutions
Table 3: Essential Research Reagents for Investigating M1 Ubiquitin Signaling
| Reagent/Category | Specific Examples | Research Application | Technical Considerations |
|---|---|---|---|
| E3 Ligase Inhibitors | HOIP/LUBAC inhibitors | Block M1 ubiquitin chain formation | Specificity for linear vs other chain types must be validated |
| Deubiquitinase Inhibitors | OTULIN, A20 modulators | Probe DUB function in signaling | May affect multiple substrates simultaneously |
| Ubiquitin Antibodies | Anti-diGly (K-ε-GG), anti-M1 linkage-specific | Ubiquitinome profiling, immunoblotting | Cross-reactivity with ubiquitin-like proteins possible |
| Cell Lines | THP1 monocytes, RAW264.7, HEK293 | Pathway manipulation studies | Species-specific differences in ubiquitin machinery |
| Animal Models | HOIP-deficient, OTULIN-deficient mice | In vivo validation of pathway function | Embryonic lethality possible with complete knockouts |
| Proteasome Inhibitors | MG132, Bortezomib | Increase ubiquitinated substrate detection | Global effects on protein turnover |
| Mass Spectrometry Standards | Heavy-labeled ubiquitin, TMT tags | Quantitative ubiquitinome comparisons | Requires specialized instrumentation and expertise |
Therapeutic Implications and Future Perspectives
The intricate balance between inflammatory signaling and cell death, regulated by ubiquitin-dependent mechanisms, presents numerous therapeutic opportunities:
Targeting Ubiquitin Pathways in Inflammatory Disease
LUBAC inhibition represents a promising approach for dampening excessive NF-κB activation in autoinflammatory disorders. The discovery that STING induces HOIP-mediated M1 ubiquitin chain synthesis suggests additional applications in type I interferonopathies [31]. Conversely, enhancing M1 ubiquitination might overcome immune suppression in certain contexts.
OTULIN-related autoinflammatory syndrome (ORAS) demonstrates the clinical relevance of linear ubiquitination regulation. The effectiveness of TNFα blockade in ORAS confirms the mechanistic connection between linear ubiquitination and TNF signaling, validating this pathway as a therapeutic target [32].
Sepsis Immunomodulation
The dual-phase nature of sepsis—initial hyperinflammation followed by immunosuppression—demands nuanced therapeutic approaches. Modulating ubiquitin-dependent cell death pathways may help rebalance the immune response:
- Apoptosis inhibition in lymphocytes could prevent sepsis-induced immunosuppression, but requires precise targeting to avoid exacerbating inflammation [36]
- Pyroptosis and NETosis inhibition may reduce tissue damage during the hyperinflammatory phase, particularly through targeting inflammasome activation and neutrophil recruitment [35]
- Ubiquitin pathway modulators that shift macrophage polarization from M1 to M2 phenotypes could facilitate inflammation resolution [32] [33]
Technical Advances and Research Directions
Future research priorities include developing higher-specificity ubiquitin reagents, including chain linkage-specific antibodies and more selective E3 ligase inhibitors. Single-cell ubiquitinome profiling would illuminate cell-to-cell heterogeneity in ubiquitin signaling during inflammation. Advanced imaging techniques to visualize M1 ubiquitin chain dynamics in live cells would provide spatial and temporal resolution currently lacking in bulk omics approaches.
The integration of ubiquitinomics with other omics datasets through systems biology approaches will further elucidate how ubiquitin networks coordinate inflammatory signaling and cell fate decisions, potentially identifying novel nodes for therapeutic intervention in inflammatory and autoimmune diseases.
Tools and Techniques: Detecting, Quantifying, and Manipulating M1-Ubiquitination in Research and Drug Discovery
Chain-Specific TUBEs (Tandem Ubiquitin Binding Entities) for High-Throughput Capture of M1-Linked Ubiquitination
The study of M1-linked, or linear, ubiquitination has emerged as a critical area in immune signaling research. This specific post-translational modification, catalyzed exclusively by the Linear Ubiquitin Chain Assembly Complex (LUBAC), serves as a key regulatory mechanism in NF-κB activation and inflammatory responses. Chain-specific Tandem Ubiquitin Binding Entities (TUBEs) have revolutionized the investigation of these signaling events by enabling high-affinity, linkage-selective capture of endogenous M1-ubiquitinated proteins. This technical guide details the application of M1-specific TUBEs for the isolation, detection, and quantification of linear ubiquitin chains in high-throughput screening formats, providing researchers with robust methodologies to advance drug discovery in inflammation and immunity.
The Linear Ubiquitination Machinery
M1-linked ubiquitination is characterized by a unique head-to-tail linkage where the C-terminal glycine of one ubiquitin molecule forms a peptide bond with the N-terminal methionine (M1) of the next ubiquitin [4] [38]. This distinctive topology differs fundamentally from the isopeptide bonds formed in lysine-linked ubiquitin chains. The synthesis of linear ubiquitin chains is exclusively catalyzed by the Linear Ubiquitin Chain Assembly Complex (LUBAC), a multi-subunit E3 ubiquitin ligase complex [4]. In humans, LUBAC consists of three core components: HOIP (HOIL-1L-Interacting Protein), which contains the catalytic RBR (RING-Between-RING) domain and linear ubiquitin chain-determining domain (LDD) essential for M1-specific chain formation; HOIL-1L (Heme-oxidized IRP2 Ubiquitin Ligase-1L); and SHARPIN (SHANK-Associated RH Domain-Interacting Protein) [4] [38]. The structural and functional integrity of all three components is required for proper regulation of linear ubiquitination in cellular signaling.
The reversibility of M1-linked ubiquitination is maintained by specialized deubiquitinases (DUBs), particularly OTULIN and CYLD, which selectively disassemble linear ubiquitin chains [4]. This precise control mechanism ensures transient and spatially regulated signaling through linear ubiquitination, preventing excessive or chronic inflammatory responses that could lead to pathological conditions.
Biological Significance in Immune Regulation
Linear ubiquitination serves as a crucial scaffolding modification in the formation of signaling complexes, particularly in the activation of the NF-κB (Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells) signaling pathway [18] [25]. Research in Drosophila melanogaster has demonstrated that the HOIP homolog LUBEL (Linear Ubiquitin E3 Ligase) catalyzes M1-ubiquitin chain formation in response to bacterial infection, which is essential for activating the Immune Deficiency (Imd) pathway and the subsequent expression of antimicrobial peptide genes [25]. This pathway shows remarkable conservation with mammalian innate immune signaling.
Beyond pathogen recognition, M1-ubiquitin chains are critical in sterile inflammation triggered by cellular stresses including hypoxia, oxidative stress, and mechanical stress [18] [39]. These chains facilitate NF-κB activation and promote cell survival under stress conditions commonly observed in chronic inflammatory diseases and cancer [18] [39]. Recent findings also implicate linear ubiquitination in STING (Stimulator of Interferon Genes) signaling, where HOIP-mediated M1-ubiquitin chain formation stimulates both NF-κB and IRF3 pathways in response to cyclic dinucleotides [31]. The broad involvement of linear ubiquitination in these diverse immune signaling cascades underscores its fundamental importance as a regulatory mechanism and potential therapeutic target.
Chain-Specific TUBE Technology
Principle and Advantages
Tandem Ubiquitin Binding Entities (TUBEs) are engineered, high-affinity reagents composed of multiple ubiquitin-associated (UBA) domains that bind polyubiquitin chains with nanomolar affinity [40] [41]. Unlike conventional antibodies, TUBEs are designed to recognize the characteristic structural features of specific ubiquitin linkage types through specialized ubiquitin-binding domains, enabling selective isolation of M1-linked chains amidst the complex landscape of cellular ubiquitination [42] [40].
The strategic advantages of TUBE technology include:
- Linkage Selectivity: M1-specific TUBEs discriminate linear ubiquitin chains from other linkage types (K48, K63, etc.) through specialized ubiquitin-binding domains that recognize the unique topology of M1-linked chains [42] [41].
- Protection from Degradation: TUBEs shield polyubiquitinated proteins from deubiquitinating enzymes (DUBs) and proteasomal degradation, preserving endogenous ubiquitination states even in the absence of protease inhibitors [41].
- High Sensitivity: With dissociation constants (Kd) in the nanomolar range (1-10 nM), TUBEs detect endogenous ubiquitination levels without requiring protein overexpression [41].
- Versatile Applications: Compatible with various downstream applications including Western blotting, mass spectrometry proteomics, high-throughput screening, and imaging [40] [41].
Comparison of Ubiquitin-Binding Reagents
Table 1: Comparison of Methods for Detecting M1-Linked Ubiquitination
| Method | Principle | Advantages | Limitations | Suitable for HTS |
|---|---|---|---|---|
| M1-Specific TUBEs | High-affinity UBA domains selective for M1 linkage | Preserves ubiquitination; nanomolar affinity; works on endogenous proteins | Requires validation of specificity | Yes [42] |
| Pan-Selective TUBEs | UBA domains binding all ubiquitin linkages | Comprehensive ubiquitin capture; high affinity | Does not discriminate linkage types | Yes [42] |
| GST-NEMO-UBAN | UBAN domain of NEMO binding linear chains | Well-characterized specificity | Lower affinity than TUBEs; requires protein expression | No [25] |
| Linkage-Specific Antibodies | Immunorecognition of linkage-specific epitopes | Wide commercial availability | Potential cross-reactivity; cannot protect from DUBs | Limited |
| Mass Spectrometry | Direct identification of linkage peptides | Definitive linkage identification | Labor-intensive; requires sophisticated instrumentation | No [42] |
Experimental Applications and Protocols
High-Throughput Capture of M1-Linked Ubiquitination
The application of chain-specific TUBEs in high-throughput screening (HTS) formats enables quantitative analysis of dynamic ubiquitination events in response to various cellular stimuli. The following workflow has been validated for investigating M1-linked ubiquitination in immune signaling contexts:
Protocol: M1-TUBE-Based Capture in 96-Well Format
Plate Coating:
- Coat 96-well plates with M1-chain selective TUBEs (e.g., LifeSensors M1 TUBEs) at a concentration of 2-5 µg/mL in PBS buffer overnight at 4°C [42].
- Block plates with 3% BSA in PBS containing 0.05% Tween-20 for 2 hours at room temperature.
Cell Stimulation and Lysis:
- Culture relevant cell types (e.g., THP-1 monocytes for immune signaling) under standard conditions.
- Stimulate cells with appropriate agonists:
- Lyse cells using a specialized buffer optimized to preserve polyubiquitination: 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% NP-40, 1 mM EDTA, 10% glycerol, supplemented with 1 mM N-ethylmaleimide (NEM) and 5 mM iodoacetamide to inhibit deubiquitinases [42]. Avoid ubiquitin-altering inhibitors when using TUBEs as they inherently protect ubiquitination.
Ubiquitin Capture and Detection:
- Incubate 50-100 µg of cell lysate per well in TUBE-coated plates for 2 hours at 4°C with gentle agitation.
- Wash plates 3-5 times with wash buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.05% Tween-20).
- Detect captured ubiquitinated proteins using:
- Immunoblotting: Use specific antibodies against proteins of interest (e.g., anti-RIPK2 for NOD2 signaling) or anti-ubiquitin antibodies [42] [43].
- Quantitative HTS Readouts: Employ horseradish peroxidase (HRP)-conjugated TUBEs or anti-ubiquitin antibodies with chemiluminescent substrates for plate readers [42].
Table 2: Experimental Conditions for Inducing M1-Linked Ubiquitination
| Signaling Context | Stimulus | Cell Models | Key Readouts | Time Course |
|---|---|---|---|---|
| NOD2/RIPK2 Signaling | L18-MDP (200-500 ng/mL) | THP-1 monocytes, BMDMs | RIPK2 ubiquitination, NF-κB activation | 30-60 minutes [42] [43] |
| Sterile Inflammation | Hypoxia (5% O₂), paraquat, mechanical stress | Drosophila, Caco-2 intestinal epithelial cells | M1-ubiquitin chain formation, cell survival, Relish/NF-κB activation | 2-24 hours [18] [39] |
| STING Signaling | Cyclic dinucleotides (cGAMP, c-di-GMP) | THP-1 monocytes, BMDMs | IRF3/NF-κB activation, M1-chain formation | 2-8 hours [31] |
| Intestinal Immunity | Oral bacterial infection (Ecc15) | Drosophila models | Antimicrobial peptide expression, pathogen clearance | 6-48 hours [25] |
Validation and Specificity Controls
Rigorous validation is essential to confirm the specificity of M1-linked ubiquitination capture:
Linkage Specificity:
Genetic Validation:
- Utilize cells with genetic ablation of LUBAC components (HOIP, HOIL-1L, SHARPIN) or CRISPR-mediated knockout to demonstrate dependence on the linear ubiquitination machinery [18] [31].
- In Drosophila models, leverage LUBEL mutant flies (lubel^Mi) to confirm specificity of M1-ubiquitin chain detection [25].
Competition Assays:
- Perform competitive inhibition with free M1-linked ubiquitin chains (0.1-1 µM) to demonstrate binding specificity.
- Use ubiquitin mutants (M1-free ubiquitin) to confirm linkage dependence.
Data Interpretation and Analysis
Quantification and Normalization Strategies
Accurate quantification of M1-linked ubiquitination signals requires appropriate normalization:
Signal Normalization:
- Normalize ubiquitination signals to total input protein levels for each sample.
- Express data as fold-change relative to unstimulated controls or time-zero timepoints.
- For target-specific ubiquitination (e.g., RIPK2), normalize ubiquitinated fraction to total target protein in input lysates [42] [43].
Kinetic Analysis:
Threshold Determination:
- Establish significance thresholds based on replicate variability (typically 2-3 fold increase over background with p<0.05).
- Define minimal detectable ubiquitination levels using positive and negative controls in each experiment.
Troubleshooting Common Technical Issues
- High Background Signal: Optimize blocking conditions, increase wash stringency, and titrate TUBE coating concentrations.
- Insufficient Signal: Verify cell stimulation efficiency, optimize lysis conditions, and confirm TUBE binding activity.
- Loss of Specificity: Include linkage-specific controls (OTULIN treatment, LUBAC knockout) in each experiment to confirm M1-specificity.
- Sample Degradation: Ensure proper inhibitor supplementation and maintain samples at 4°C during processing.
Research Reagent Solutions
Table 3: Essential Reagents for M1-Linked Ubiquitination Research
| Reagent | Specific Example | Function/Application | Commercial Source/Reference |
|---|---|---|---|
| M1-Specific TUBEs | M1-TUBE (UM401M series) | Selective capture of linear ubiquitin chains | LifeSensors [41] |
| Pan-Selective TUBEs | TUBE2 (UM202) | Comprehensive ubiquitin capture for comparison | LifeSensors [40] [41] |
| LUBAC Inhibitors | HOIPIN-8 | Specific inhibition of LUBAC catalytic activity | Literature compounds [4] |
| DUBs for Specificity Control | Recombinant OTULIN | Selective cleavage of M1-linked chains for validation | Commercial vendors [25] [4] |
| Positive Control Stimuli | L18-MDP | Induces K63/M1 ubiquitination in NOD2/RIPK2 pathway | Commercial vendors [42] [43] |
| LUBAC Components | HOIP, HOIL-1L, SHARPIN antibodies | Validation of LUBAC expression and integrity | Multiple commercial sources |
| Specialized Lysis Buffer | DUB-inhibiting buffer | Preserves endogenous ubiquitination states | Custom formulation [42] |
Signaling Pathway Visualization
Figure 1: M1-Linked Ubiquitination in Immune Signaling Pathways. This diagram illustrates the central role of linear ubiquitin chains in multiple innate immune signaling pathways. The LUBAC complex generates M1-linked ubiquitin chains in response to various stimuli, including pathogen-associated molecular patterns (PAMPs), damage-associated molecular patterns (DAMPs), and cyclic dinucleotides. These chains facilitate the activation of key signaling complexes, particularly the IKK complex, leading to NF-κB and IRF3 activation and subsequent inflammatory gene expression.
Chain-specific TUBEs represent a transformative technology for deciphering the complex roles of M1-linked ubiquitination in immune signaling. The methodologies outlined in this technical guide provide researchers with robust, reproducible tools to investigate linear ubiquitination dynamics in high-throughput formats, enabling accelerated discovery in both basic immune mechanism research and therapeutic development. As our understanding of the ubiquitin code continues to evolve, TUBE-based approaches will remain essential for elucidating the precise functions of M1-linked ubiquitination in health and disease.
Mass Spectrometry-Based Proteomics for Mapping Linear Ubiquitination Sites
Linear ubiquitination, also known as M1-linked ubiquitination, is a distinctive post-translational modification (PTM) characterized by a head-to-tail linkage of ubiquitin molecules through a peptide bond between the C-terminal glycine of the donor ubiquitin and the N-terminal methionine of the acceptor ubiquitin [38]. Unlike other ubiquitin linkage types that form isopeptide bonds via lysine residues, this unique linear architecture is exclusively catalyzed by the Linear Ubiquitin Chain Assembly Complex (LUBAC), a multi-protein complex comprising HOIP, HOIL-1, and SHARPIN [44] [38]. LUBAC-mediated linear ubiquitination has emerged as a critical regulator of immune signaling pathways, including NF-κB activation, immune cell development, and cell death regulation, with implications for inflammatory diseases and cancer [44].
The detection and mapping of linear ubiquitination sites present substantial technical challenges due to the low stoichiometry of endogenous ubiquitination, the transient nature of signaling events, and the complexity of the ubiquitin code [45] [37]. Mass spectrometry (MS)-based proteomics has become the cornerstone technology for comprehensively profiling protein ubiquitination, enabling researchers to identify substrates, map modification sites, and characterize chain topology with unprecedented precision [45] [46]. This technical guide examines current methodologies, experimental workflows, and analytical strategies for mapping linear ubiquitination sites, with particular emphasis on their application to immune signaling research.
Technical Foundations of Ubiquitin Proteomics
The Unique Molecular Architecture of Linear Ubiquitination
The linear ubiquitination machinery consists of specialized enzymes that write, read, and erase this modification. LUBAC serves as the sole E3 ligase complex capable of generating M1-linked chains in vertebrates [38]. HOIP contains the catalytic RING-IBR-RING (RBR) domain and a unique C-terminal linear ubiquitin chain determining domain (LDD) that positions the acceptor ubiquitin for M1-linkage formation [38]. The complex is regulated through autoinhibitory mechanisms and requires specific interactions between the UBA domain of HOIP and the UBL domains of HOIL-1 and SHARPIN for full activation [38].
The linear ubiquitin signal is decoded by specific reader proteins containing ubiquitin-binding domains (UBDs) with preference for M1-linkages, such as NF-κB essential modulator (NEMO), which plays crucial roles in inflammatory signaling [38]. Deubiquitinases (DUBs) including OTULIN and CYLD function as erasers that specifically disassemble linear chains, ensuring the transient nature of this modification in signaling events [44] [38]. This precise regulation underscores the importance of linear ubiquitination in physiological processes and highlights the need for specific detection methods.
Mass Spectrometry-Based Approaches for Ubiquitinome Analysis
Mass spectrometry has revolutionized the study of protein ubiquitination by enabling large-scale identification and quantification of modification sites. The diGly remnant approach represents the most widely used strategy, leveraging the fact that tryptic digestion of ubiquitinated proteins leaves a characteristic di-glycine (diGly) signature on modified lysine residues with a detectable mass shift of 114.04 Da [45] [46]. This approach has been successfully applied to profile linear ubiquitination sites despite the unique linkage type, as the C-terminal Gly76 of ubiquitin forms the isopeptide bond with the substrate lysine regardless of chain topology [37].
Table 1: Mass Spectrometry Acquisition Methods for Ubiquitinome Analysis
| Method | Principle | Advantages | Limitations | Typical Identifications (diGly sites) |
|---|---|---|---|---|
| Data-Dependent Acquisition (DDA) | Intensity-based selection of top precursors for fragmentation | Well-established, comprehensive spectral libraries | Missing values across samples, limited dynamic range | ~20,000 in single runs [37] |
| Data-Independent Acquisition (DIA) | Parallel fragmentation of all ions in predefined m/z windows | Higher sensitivity, better quantitative accuracy, fewer missing values | Requires extensive spectral libraries | ~35,000 in single runs [37] |
| Label-Free Quantification | Direct comparison of precursor intensities across runs | Cost-effective, no chemical labeling required | Higher technical variability | Suitable for large sample cohorts [46] |
| Isobaric Labeling (TMT, SILAC) | Incorporation of stable isotopes for multiplexing | High multiplexing capacity, reduced missing values | Ratio compression effects | Enhanced throughput for condition comparisons [46] |
Recent advancements in Data-Independent Acquisition (DIA) methods have dramatically improved the sensitivity and reproducibility of ubiquitinome analyses. A 2021 study demonstrated that DIA nearly doubles the number of diGly peptide identifications in single measurements compared to Data-Dependent Acquisition (DDA), with approximately 35,000 distinct diGly sites identified from proteasome inhibitor-treated cells [37]. This enhanced coverage is crucial for capturing the low-abundance linear ubiquitination events that govern immune signaling pathways.
Experimental Workflows for Linear Ubiquitination Mapping
Sample Preparation and Enrichment Strategies
Effective enrichment of ubiquitinated peptides is essential for comprehensive linear ubiquitination mapping due to the low stoichiometry of this modification. The following section outlines key methodologies for sample preparation and enrichment.
Figure 1: Experimental Workflow for Linear Ubiquitination Site Mapping
Antibody-Based Enrichment Methods
Immunoaffinity purification using anti-diGly remnant antibodies represents the gold standard for ubiquitinome studies. This approach specifically enriches peptides containing the K-ε-GG motif after tryptic digestion, capturing all ubiquitin linkage types including linear chains [46] [37]. Optimization of antibody-to-peptide ratios is critical for maximizing recovery; recent studies indicate that 31.25 μg of anti-diGly antibody per 1 mg of peptide input provides optimal enrichment efficiency [37]. For specialized applications, linkage-specific antibodies targeting M1-linked chains can be employed, though these are less commonly used for proteome-wide studies due to availability and specificity constraints [45].
Alternative Enrichment Strategies
Affinity tag-based purification utilizes cells expressing epitope-tagged ubiquitin (e.g., His, FLAG, or Strep tags) for purification under denaturing conditions. The tagged ubiquitin system enables purification of ubiquitinated proteins before digestion, reducing sample complexity [45]. However, this approach requires genetic manipulation and may not fully recapitulate endogenous regulation.
Ubiquitin-binding domain (UBD)-based enrichment leverages natural ubiquitin receptors such as tandem-repeated UBD domains with enhanced affinity for polyubiquitin chains. While this method preserves endogenous modification patterns, it typically exhibits lower specificity compared to antibody-based approaches [45].
Mass Spectrometry Analysis and Data Acquisition
Following enrichment, diGly-modified peptides are analyzed using high-resolution mass spectrometry. The unique properties of diGly peptides—including longer sequence lengths and higher charge states due to impeded C-terminal cleavage at modified lysines—require specialized MS method optimization [37].
Optimal DIA parameters for diGly peptide analysis include:
- Precursor isolation windows: 46 windows with optimized widths based on precursor distribution
- MS2 resolution: 30,000 for improved signal-to-noise ratio
- Cycle time: Balanced to ensure sufficient chromatographic sampling [37]
For spectral library generation, deep reference maps are constructed by fractionating diGly-enriched peptides (typically 8-96 fractions) followed by DDA analysis. Combining libraries from multiple cell lines and conditions significantly increases coverage; recent studies have assembled libraries containing >90,000 diGly peptides [37]. These comprehensive resources enable robust identification and quantification of linear ubiquitination sites across diverse biological contexts.
Advanced Applications in Immune Signaling Research
Systematic Interactome Profiling of LUBAC Components
Comprehensive profiling of HOIP interactomes across tissues has revealed novel insights into linear ubiquitination functions in immune homeostasis. A 2024 study employed affinity purification mass spectrometry (AP-MS) with four HOIP truncations (PUB, NZF, UBA, and RBR-LDD domains) across nine mouse tissues, identifying an average of 90 interactors per bait in each tissue [44]. This systematic approach uncovered tissue-specific interaction patterns for known HOIP partners; for instance, STAT1 showed strong interaction with HOIP in liver, lung, and colorectal tissues, while β-Catenin interactions were prominent in brain, liver, lung, stomach, and colorectal tissues [44].
Table 2: Key HOIP Domain Functions and Substrates in Immune Signaling
| HOIP Domain | Primary Functions | Key Interacting Partners | Role in Immune Signaling |
|---|---|---|---|
| PUB | OTULIN and SPATA2 binding, deubiquitinase regulation | OTULIN, CYLD, SPATA2 | Negative regulation of linear ubiquitination, DUB recruitment [44] |
| NZF | Substrate recognition, ubiquitin chain binding | NEMO, STAT1, ALK1 | Substrate selection for linear ubiquitination [44] |
| UBA | Complex stabilization, HOIL-1 and SHARPIN recruitment | HOIL-1, SHARPIN | LUBAC assembly and activation [44] [38] |
| RBR-LDD | Catalytic activity, linear chain formation | E2 ubiquitin-conjugating enzymes | M1-linked ubiquitin chain synthesis [44] [38] |
This domain-resolved interactome mapping identified novel linear ubiquitination substrates with implications for immune function, including Integrin-linked kinase (ILK) and Smad ubiquitination regulatory factor 1 (SMURF1) [44]. Linear ubiquitination of ILK was shown to decrease focal adhesion formation and promote detachment of Shigella flexneri-infected cells, illustrating how pathogens manipulate host linear ubiquitination for immune evasion [44].
Quantitative Dynamics in Signaling Pathways
Advanced quantitative proteomics has enabled time-resolved analysis of linear ubiquitination dynamics during immune activation. Applying DIA-based diGly proteomics to TNFα signaling—a pathway critically dependent on LUBAC-mediated linear ubiquitination—researchers comprehensively captured known regulatory sites while identifying numerous novel modifications [37]. The enhanced quantitative accuracy of DIA (45% of diGly peptides with CVs <20% compared to 15% with DDA) proved particularly valuable for capturing transient phosphorylation events that coordinate with linear ubiquitination in NF-κB activation [37].
Circadian regulation of ubiquitination represents another emerging application, with systems-wide analyses revealing hundreds of cycling ubiquitination sites and clusters within membrane protein receptors and transporters [37]. These findings highlight new connections between ubiquitination dynamics and circadian biology, with potential implications for immune function given the established links between circadian rhythms and immune responses.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Linear Ubiquitination Studies
| Reagent Category | Specific Examples | Applications | Considerations |
|---|---|---|---|
| Anti-diGly Antibodies | PTMScan Ubiquitin Remnant Motif Kit (CST) [37] | Immunoaffinity enrichment of diGly peptides | Commercial antibodies optimized for K-ε-GG motif enrichment |
| Linkage-Specific Antibodies | M1-linkage specific antibodies [45] | Selective enrichment of linear ubiquitin chains | Limited availability, require validation for specific applications |
| Tagged Ubiquitin Systems | His-tagged Ub, Strep-tagged Ub [45] | Affinity purification of ubiquitinated proteins | May not fully replicate endogenous regulation |
| LUBAC Components | Recombinant HOIP truncations (PUB, NZF, UBA, RBR-LDD) [44] | Interaction studies, in vitro ubiquitination assays | Domain-specific functions require careful experimental design |
| Deubiquitinases | Recombinant OTULIN, CYLD [44] [38] | Specific cleavage of linear ubiquitin chains | Tool for validating M1-linkage specificity |
| Mass Spectrometry Standards | Heavy-labeled reference peptides | Quantitative accuracy assessment | Essential for absolute quantification studies |
Future Perspectives and Concluding Remarks
The evolving methodology for mapping linear ubiquitination sites continues to reshape our understanding of immune signaling mechanisms. Recent technical innovations—particularly in DIA-based acquisition and enrichment strategies—have dramatically improved the sensitivity, coverage, and quantitative accuracy of ubiquitinome studies [37]. These advances are enabling researchers to capture the dynamic and transient nature of linear ubiquitination in immune activation with unprecedented temporal resolution.
Future methodological developments will likely focus on improving specificity for M1-linked chains within complex ubiquitin landscapes, enhancing spatial resolution through subcellular fractionation or proximity labeling, and integrating multi-omics approaches to understand cross-talk between linear ubiquitination and other PTMs. Additionally, the application of targeted proteomics methods like parallel reaction monitoring (PRM) could provide validated assays for specific linear ubiquitination events of high biological significance [46].
As these methodologies continue to mature, mass spectrometry-based proteomics will remain an indispensable tool for deciphering the complex roles of linear ubiquitination in immune signaling, with broad implications for understanding disease mechanisms and developing targeted therapeutic interventions. The systematic application of these approaches across diverse immune cell types and activation states promises to uncover novel regulatory mechanisms and expand our understanding of how linear ubiquitination shapes immune responses.
The post-translational modification of proteins with ubiquitin is a sophisticated mechanism for regulating cellular signalling systems. Among the different types of ubiquitin chains, Met1-linked (or linear) ubiquitin has emerged as a key regulatory signal that controls cell death, immune signalling, and other vital cellular functions [47]. The Linear Ubiquitin Chain Assembly Complex (LUBAC), the only known E3 ubiquitin ligase in vertebrates that specifically assembles Met1-linked ubiquitin chains, plays essential roles in regulating NF-κB activation, immune responses, and inflammation [38] [4]. LUBAC is composed of three core subunits: HOIP (RNF31), HOIL-1 (RBCK1), and SHARPIN [47]. HOIP serves as the catalytic core of the complex, containing a unique C-terminal linear ubiquitin chain determining domain (LDD) that positions the N-terminus of the acceptor ubiquitin for Met1-linkage formation [38]. Both HOIL-1 and SHARPIN play crucial structural roles in stabilizing the complex and relieving the autoinhibition of HOIP's catalytic activity [38].
Understanding LUBAC functions requires precise genetic manipulation strategies to modulate its components and the ubiquitin system itself. This technical guide provides an in-depth overview of current methodologies for LUBAC component knockout and mutant ubiquitin expression, framed within the context of investigating M1-linear ubiquitin chain functions in immune signalling research. These approaches have revealed that LUBAC modulates CBM complex functions downstream of TRAF6 in T cells, influences NLRP3 inflammasome activation in macrophages, and regulates both cell death and survival pathways in a context-dependent manner [9] [48] [4].
Genetic Knockout Strategies for LUBAC Components
CRISPR/Cas9-Mediated Knockout in Primary Human T Cells
Knocking out LUBAC components in primary human immune cells provides critical insights into cell-type-specific functions of linear ubiquitination. The following protocol has been successfully employed to generate knockout pools in primary human CD4+ T cells [9]:
Key Technical Considerations:
- Optimization: Initial optimization using sgRNA targeting TCRα achieved ~50% homozygous knockout efficiency as measured by loss of TCRα/β surface expression [9].
- Efficiency: With sgRNAs targeting LUBAC components, approximately 85% reduction of HOIP protein and 60% reduction of TRAF6 protein was achieved in pooled populations [9].
- Validation: Western blot analysis confirmed that targeting HOIP or HOIL-1 causes LUBAC instability, with loss of both subunits occurring when either is targeted due to complex destabilization [9].
Cell-Type Specific Phenotypes of LUBAC Knockouts
Genetic ablation of LUBAC components produces distinct phenotypes across different cell types, highlighting the context-dependent functions of linear ubiquitination:
| Cell Type | Genetic Manipulation | Key Phenotypes | Functional Assessment |
|---|---|---|---|
| Primary human CD4+ T cells | HOIP/HOIL-1 KO | Mild impairment of TCR-induced NF-κB; altered MALT1 substrate recognition | IκBα degradation, p65 phosphorylation, MALT1 substrate cleavage [9] |
| Bone marrow-derived macrophages | HOIL-1 KO | Normal IκBα degradation; defective NLRP3 inflammasome activation | IL-1β secretion, ASC oligomerization, caspase-1 cleavage [48] |
| Mouse embryonic fibroblasts | HOIL-1 KO | Delayed IκBα degradation; reduced NF-κB activation | IκBα degradation kinetics, IL-6 secretion [48] |
| Jurkat T cells | HOIP KO | Reduced CBM-dependent NF-κB activation | NF-κB reporter assays, target gene expression [9] |
Mutant Ubiquitin Expression Strategies
Tetracycline-Inducible Ubiquitin Replacement System
To investigate the specific roles of different ubiquitin linkage types, researchers have developed a sophisticated ubiquitin replacement methodology that enables endogenous ubiquitin knockdown with simultaneous expression of mutant ubiquitin [49]. This system is particularly valuable for studying the functional requirements of specific ubiquitin linkages in signalling pathways.
Experimental Workflow:
- Design shRNA sequences targeting all four endogenous ubiquitin genes:
- sh-Ub1: Targets UBC and UBA52
- sh-Ub2: Targets UBB and RPS27A
- Generate stable cell lines with tetracycline-inducible shRNA constructs
- Create rescue constructs with RNAi-resistant wild-type or mutant ubiquitin (e.g., K63R)
- Select stable clones capable of inducible ubiquitin replacement
- Validate replacement efficiency via RT-PCR and immunoblotting
This approach achieved 80-95% reduction of endogenous ubiquitin transcripts with simultaneous expression of mutant ubiquitin variants [49].
Commonly Used Ubiquitin Mutants in Linear Ubiquitination Research
| Ubiquitin Mutant | Key Properties | Applications in Signaling Research |
|---|---|---|
| K63R | Cannot form K63-linked chains; essential for IL-1β but not TNFα-induced IKK activation | Dissecting K63 vs linear ubiquitin requirements [49] |
| M1-only (all lysines mutated) | Permits only linear ubiquitination | Studying exclusive functions of M1-linked chains |
| K48R | Defective in proteasomal targeting | Distinguishing proteolytic vs non-proteolytic functions |
| K11R | Disrupts K11-linked branching | Investigating branched chain functions in proteasomal recognition [50] |
Experimental Protocols for Functional Validation
Assessing NF-κB Signalling in LUBAC-Modified Cells
Protocol: Time-Course Analysis of NF-κB Activation
- Stimulation: Treat control and LUBAC-deficient cells with PMA/lonomycin (T cells) or TNFα/LPS (other cell types) for 0, 15, 30, 60, and 120 minutes [9].
- Cell Lysis: Harvest cells in RIPA buffer supplemented with protease and phosphatase inhibitors.
- Western Blot Analysis: Probe for:
- IκBα degradation (early marker of NF-κB activation)
- Phospho-p65 (Ser536) as activation marker
- Total p65 as loading control
- Single-Cell Analysis: For primary T cells, perform intracellular staining for IκBα and phospho-p65 followed by flow cytometry to account for heterogeneous knockout efficiency [9].
- Nuclear Translocation: Use imaging flow cytometry to quantify p65 nuclear accumulation [9].
Analysis of MALT1 Protease Activity
Protocol: MALT1 Substrate Cleavage Assay
- Cell Stimulation: Activate T cells via TCR engagement (anti-CD3/CD28) or PMA/lonomycin for 2-24 hours [9].
- Protein Extraction: Prepare whole cell lysates in appropriate buffer.
- Western Blot Detection: Probe for cleavage of known MALT1 substrates:
- HOIL-1 (full length and cleavage fragment)
- CYLD
- A20
- Regnase-1
- Densitometric Analysis: Quantify the ratio of cleaved to full-length substrates.
Research Reagent Solutions
Essential tools and reagents for genetic manipulation of the linear ubiquitination system:
| Research Tool | Specifications | Key Applications |
|---|---|---|
| LUBAC subunit sgRNAs | CRISPR/Cas9 guides targeting HOIP, HOIL-1, SHARPIN | Generation of LUBAC-deficient cells [9] |
| Tetracycline-inducible ubiquitin replacement system | shRNA-resistant ubiquitin mutants | Linkage-specific ubiquitin function studies [49] |
| Linear ubiquitin-specific antibodies | Anti-M1 linkage antibodies | Detection of endogenous linear ubiquitination [38] |
| OTULIN and CYLD constructs | Catalytically active and inactive mutants | Manipulating linear ubiquitin chain disassembly [4] |
| LUBAC complex recombinant proteins | HOIP/HOIL-1/SHARPIN trimeric complex | In vitro ubiquitination assays [38] |
Analysis of Signaling Pathways in LUBAC-Modified Cells
Understanding how genetic manipulations of LUBAC components affect immune signalling pathways requires comprehensive pathway analysis. The diagrams below illustrate key pathways and their modulation by LUBAC.
LUBAC in T Cell Receptor Signalling
Experimental Workflow for Comprehensive LUBAC Analysis
Genetic manipulation of LUBAC components and the ubiquitin system provides powerful approaches to dissect the specific functions of linear ubiquitination in immune signalling. The strategies outlined in this technical guide – from CRISPR-mediated knockout in primary cells to sophisticated ubiquitin replacement systems – enable researchers to probe the complex roles of linear ubiquitination in NF-κB activation, inflammasome regulation, and cell death pathways. The experimental protocols and reagent solutions detailed here provide a foundation for designing comprehensive studies to address specific research questions in this rapidly evolving field. As research progresses, these methodologies will continue to reveal the intricate mechanisms through which linear ubiquitination governs immune homeostasis and contributes to human disease pathologies.
The functional assessment of nuclear factor kappa B (NF-κB) signaling and concomitant cytokine responses represents a cornerstone of immunological research, particularly in the context of innate immunity and inflammation. Within this signaling landscape, M1-linked or linear ubiquitin chains have emerged as crucial post-translational modifications that exert sophisticated control over immune activation pathways. These specialized ubiquitin chains, formed through head-to-tail linkage via the N-terminal methionine of ubiquitin, serve as specific regulatory signals that control cell death, immune activation, and inflammation [47] [38]. The Linear Ubiquitin Chain Assembly Complex (LUBAC), comprised of HOIP, HOIL-1L, and SHARPIN, stands as the sole E3 ubiquitin ligase capable of generating these M1-linked ubiquitin chains, while dedicated deubiquitinases (DUBs) such as OTULIN and CYLD provide stringent negative regulation [4].
This technical guide provides researchers with comprehensive methodologies for interrogating NF-κB activation and cytokine responses within experimental systems where M1 ubiquitination is manipulated or monitored. The integration of these techniques enables a multi-layered functional analysis of immune signaling pathways, offering critical insights for both basic research and drug discovery applications targeting inflammatory diseases, cancer, and immune disorders.
Technical Foundation: M1 Ubiquitin Chain Biology and NF-κB Signaling
The Linear Ubiquitination Machinery
The enzymatic machinery governing M1-linked ubiquitin chains operates through a highly specific system of writers, readers, and erasers:
LUBAC Complex: HOIP serves as the catalytic core containing a unique C-terminal linear ubiquitin chain determining domain (LDD) that positions the N-terminus of the target ubiquitin for M1-linkage formation. HOIL-1L and SHARPIN function as essential adaptors that stabilize the complex and relieve HOIP's autoinhibition [38] [4]. The interaction between the ubiquitin-associated (UBA) domain of HOIP and ubiquitin-like (UBL) domains of HOIL-1L and SHARPIN maintains complex integrity [4].
Regulatory DUBs: OTULIN (OTU deubiquitinase with linear linkage specificity) demonstrates remarkable specificity for cleaving M1-linked ubiquitin chains through its ovarian tumor (OUT) domain containing conserved catalytic residues (Cys129, His339, Asn341) [4]. CYLD (cylindromatosis gene) also disassembles linear ubiquitin chains, though with broader specificity that includes K63-linked chains [4].
M1 Ubiquitination in Immune Pathway Activation
Linear ubiquitin chains function as scaffolds for protein complex assembly rather than signals for proteasomal degradation. Recent research has illuminated how this specialized ubiquitination controls NF-κB signaling:
STING Pathway Integration: Recent findings demonstrate that STING activation induces HOIP-mediated synthesis of M1 ubiquitin chains to stimulate NF-κB signaling. Upon cyclic dinucleotide binding, STING recruits LUBAC to LC3B-associated Golgi membranes where M1-ubiquitin chain formation occurs, establishing a novel mechanism for STING-induced NF-κB activation independent of its role in IRF3 signaling [31].
TNFR and IL-1R Signaling: LUBAC-generated linear ubiquitin chains modify components of the TNF receptor and IL-1 receptor signaling complexes, facilitating NEMO binding and subsequent IKK complex activation, which ultimately phosphorylates IκBα leading to its degradation and NF-κB nuclear translocation [47] [4].
Feedback Regulation: Evidence from zebrafish injury models reveals an NF-κB/TNFα signaling feedback loop where NF-κB activity modulates TNFα expression to control macrophage migration and inflammatory responses during tissue regeneration [51].
Figure 1: M1 Linear Ubiquitin-Dependent NF-κB Activation Pathways. Linear ubiquitin chains assembled by LUBAC serve as critical signaling scaffolds in multiple NF-κB activation pathways, including cytokine receptor signaling and the recently discovered STING pathway that recruits LUBAC to Golgi membranes. Regulatory DUBs (OTULIN/CYLD) provide negative feedback by disassembling M1 linkages.
Experimental Approaches: Reporter Assays and Cytokine Profiling
NF-κB Reporter Assays: Methodological Framework
NF-κB reporter assays provide a direct measurement of pathway activity by quantifying transcription factor-dependent expression of easily measurable reporter genes.
Reporter Construct Design and Validation
- Luciferase-Based Reporters: The most common approach utilizes firefly or NanoLuc luciferase genes under control of NF-κB response elements, typically comprising multiple copies of the consensus GGGRNNYYCC sequence (where R = purine, Y = pyrimidine, N = any nucleotide) [52] [53].
- Dual-Luciferase Systems: Experimental configurations should incorporate Renilla luciferase under constitutive promoters (e.g., CMV, SV40) for normalization against transfection efficiency and cellular viability [52].
- Oscillatory Dynamics Analysis: Recent advances employ live-cell luciferase systems to monitor NF-κB oscillatory profiles over time, which can decode differential responses to therapeutic agents like cetuximab in inflammatory environments [53].
Detailed Protocol: Dual-Luciferase NF-κB Reporter Assay
Table 1: Key Reagents for NF-κB Reporter Assays
| Reagent/Cell Line | Specifications | Experimental Function |
|---|---|---|
| Reporter Construct | pNF-κB-Luc (Firefly), pRL-TK (Renilla) | Primary reporter and normalization control |
| Stimuli | LPS (1 μg/mL), TNF-α (10-50 ng/mL), IL-1β (10-20 ng/mL) | Pathway activation [52] |
| LUBAC Modulators | MLKL-IN1 (LUBAC inhibitor), HOIP overexpression plasmids | Specific manipulation of linear ubiquitination |
| Cell Lines | HEK293T, THP-1 monocytes, Mouse BMDMs | Model systems for signaling studies [31] |
| Inhibitors | BMS-345541 (IKK2 inhibitor, 2.5 mM), JSH-23 (p65 nuclear translocation inhibitor, 300 μM) | Pathway inhibition controls [51] |
| Detection Reagents | Dual-Luciferase Reporter Assay System | Luminescence quantification |
Day 1: Cell Seeding and Transfection
- Seed appropriate cell line (e.g., HEK293T at 1.5×10^5 cells/well in 24-well plates) in complete medium without antibiotics.
- Transfect cells once adhered (typically 70-90% confluence) using preferred transfection reagent with the following DNA mixture per well:
- 400 ng pNF-κB-Luc reporter plasmid
- 40 ng pRL-TK normalization plasmid
- 100-500 ng experimental plasmids (LUBAC components, mutants, or empty vector control)
- Incubate transfected cells for 24-48 hours to allow reporter expression.
Day 2/3: Stimulation and Pathway Modulation
- Pre-treat cells with experimental modulators (LUBAC inhibitors, cytokine antagonists) 1-2 hours prior to stimulation.
- Stimulate NF-κB pathway with appropriate agonists:
- LPS at 1 μg/mL for TLR4 activation [52]
- TNF-α at 10-50 ng/mL for TNFR activation
- IL-1β at 10-20 ng/mL for IL-1R activation
- Incubate for predetermined timepoints (typically 4-8 hours for peak response).
Day 2/3: Luciferase Measurement
- Aspirate medium and lyse cells with 100 μL 1× Passive Lysis Buffer per well.
- Transfer lysates to microcentrifuge tubes, vortex briefly, and centrifuge at 12,000×g for 1 minute.
- Program luminometer for dual-luciferase measurement with 2-second measurement intervals.
- For each sample, aliquot 20 μL lysate into luminometer tube, inject 100 μL Luciferase Assay Reagent II, measure firefly luminescence.
- Quench firefly reaction and activate Renilla luciferase by injecting 100 μL Stop & Glo Reagent, measure Renilla luminescence.
- Calculate normalized NF-κB activity as Firefly Luciferase/Renilla Luciferase for each sample.
Advanced Applications: Live-Cell Kinetic Profiling
For oscillatory dynamics assessment [53]:
- Seed cells stably expressing NF-κB-luciferase reporters in 96-well black-walled plates.
- Equilibrate with luciferin substrate (150 μg/mL final concentration) in imaging medium.
- Record baseline luminescence for 30-60 minutes before stimulation.
- Automatically inject stimuli and continue recording for 12-24 hours with 5-15 minute intervals.
- Analyze oscillation frequency, amplitude, and duration using specialized software (e.g., Matlab algorithms for rhythm analysis).
Cytokine Profiling: Multiplexed Approaches
Comprehensive cytokine analysis provides functional readouts of NF-κB activation downstream of M1 ubiquitin signaling, capturing the physiological output of pathway activity.
Method Selection Matrix
Table 2: Cytokine Profiling Methodologies and Applications
| Methodology | Sensitivity Range | Multiplexing Capacity | Sample Throughput | Key Applications |
|---|---|---|---|---|
| Multiplex Bead Arrays (Luminex) | 1-10 pg/mL | 17-100 analytes | Medium-High | Comprehensive immune monitoring [54] |
| Single-Cell RNA Sequencing | Transcript-level | Genome-wide | Low-Medium | Cell-type specific responses [55] |
| ELISA | 1-10 pg/mL | Single analyte | High | Targeted, high-precision quantification |
| MSD Electrochemiluminescence | 0.1-1 pg/mL | 10-40 analytes | High | High-sensitivity validation studies [54] |
Detailed Protocol: Multiplex Cytokine Analysis of Primary Immune Cells
PBMC Isolation and Stimulation
- Collect peripheral blood in sodium heparin or EDTA tubes, dilute 1:1 with PBS.
- Isolate PBMCs using Ficoll-Paque density gradient centrifugation:
- Layer diluted blood carefully over Ficoll (3:2 blood:Ficoll ratio)
- Centrifuge at 400×g for 30 minutes at 20°C with brake disengaged
- Collect PBMC interface, wash twice with PBS (250×g, 10 minutes)
- Count cells and resuspend at 2×10^6 cells/mL in complete RPMI 1640.
- Plate 1 mL cell suspension per well in 24-well plates.
- Stimulate with pathway-specific agonists:
- αCD3/αCD28 beads (1:1 bead:cell ratio) for T-cell activation
- LPS (100 ng/mL) for myeloid cell activation
- PHA (5 μg/mL) for broad lymphocyte activation
- SEB/SEA (10 ng/mL each) for superantigen response [54]
- Incubate for 48 hours at 37°C, 5% CO₂.
Cytokine Measurement and Data Analysis
- Collect supernatant by centrifugation at 500×g for 5 minutes.
- Aliquot and store at -80°C until analysis.
- Thaw samples on ice and analyze using customized multiplex platforms (Luminex, MSD):
- Include standard curves for absolute quantification
- Incorporate quality controls for inter-assay normalization
- Measure key NF-κB-dependent cytokines:
- Pro-inflammatory: IL-1β, IL-6, IL-8, TNF-α
- Regulatory: IL-10
- Adaptive interface: IL-12, IL-23
- Normalize data to cell number or constitutive markers.
- Utilize specialized analysis approaches for single-cell data:
Figure 2: Integrated Experimental Workflow for NF-κB and Cytokine Analysis. The parallel implementation of NF-κB reporter assays and cytokine profiling creates complementary datasets that, when integrated, provide comprehensive insights into how M1 ubiquitin chains regulate immune signaling from initial pathway activation to functional cellular outputs.
Data Interpretation and Integration
Analytical Considerations for M1 Ubiquitin Studies
Temporal Dynamics: NF-κB activation demonstrates oscillatory behavior with important functional implications. Treatments that alter oscillation frequency or duration may indicate specific M1 ubiquitin-dependent regulation mechanisms [53].
Cell-Type Specific Responses: Cytokine responses exhibit remarkable cell-type specificity. For example, IL-1β induces distinct gene programs in different immune cell types, which may be differentially regulated by linear ubiquitination [55].
Feedback Loop Integration: Experimental designs must account for inherent feedback regulation, such as the NF-κB/TNFα feedback loop where NF-κB inhibition unexpectedly increases TNFα expression and enhances macrophage migration to injury sites [51].
Troubleshooting Common Technical Challenges
Reporter Assay Variability: Excessive variability in normalized luciferase activity often stems from transfection efficiency differences. Solutions include:
- Implementing stable cell lines with integrated reporters
- Using baculoviral or lentiviral delivery systems for primary cells
- Extending incubation time post-transfection to 48 hours
Cytokine Detection Sensitivity: Suboptimal detection of low-abundance cytokines can be addressed by:
- Concentrating supernatant samples using centrifugal filters
- Switching to higher-sensitivity platforms (MSD electrochemiluminescence)
- Pre-treatment with protein transport inhibitors for intracellular cytokine staining
M1 Ubiquitin Specificity: Ensuring observed effects specifically relate to linear ubiquitination requires:
- Multiple complementary approaches (genetic LUBAC disruption, OTULIN overexpression)
- Reciprocal experiments with catalytic LUBAC mutants
- Validation with linkage-specific ubiquitin binding domains
The synergistic application of NF-κB reporter assays and cytokine profiling provides a powerful methodological framework for deciphering how M1 linear ubiquitin chains control immune signaling outcomes. These techniques enable researchers to move beyond static protein measurements to capture dynamic signaling events and functional cellular responses. As the role of linear ubiquitination expands beyond canonical NF-κB activation to include emerging areas such as STING signaling [31], cell death regulation [4], and tissue regeneration [51], these assay approaches will continue to provide critical insights into immune mechanism and therapeutic opportunities. The integration of advanced methodologies—particularly single-cell transcriptomics and live-cell kinetic profiling—promises to further illuminate the sophisticated regulatory functions of this specialized ubiquitin code in health and disease.
The post-translational modification of proteins by M1-linked, or linear, ubiquitin chains is a crucial regulatory mechanism in innate and adaptive immune signaling. Unlike other ubiquitin chain types that form via lysine residues, linear chains are formed by the Linear Ubiquitin Chain Assembly Complex (LUBAC) through a peptide bond between the C-terminal glycine of one ubiquitin and the N-terminal methionine (M1) of another [38] [4]. This specific modification serves as a critical scaffold for the assembly of signaling complexes, particularly in the activation of the canonical NF-κB pathway, a master regulator of inflammation and immune responses [56] [4]. The activity of LUBAC is antagonized by the deubiquitinase OTULIN, the only enzyme known to specifically cleave linear ubiquitin chains [56] [4]. The precise balance between LUBAC-mediated linear ubiquitination and OTULIN-mediated deubiquitination is therefore a fundamental checkpoint in immune homeostasis, and its dysregulation is implicated in cancer, autoinflammatory disorders, and neurodegenerative diseases [57] [58] [59]. This whitepaper provides an in-depth technical guide for drug discovery campaigns aimed at identifying and characterizing pharmacological modulators of these key enzymes.
Biological Background and Therapeutic Rationale
The Linear Ubiquitination Machinery: Writers, Erasers, and Readers
The core machinery for linear ubiquitination is composed of a minimal set of highly specific components.
LUBAC (The Writer): LUBAC is a multi-subunit E3 ubiquitin ligase and the only known enzyme capable of forming linear ubiquitin chains [4]. Its catalytic core consists of three proteins: HOIP (RNF31), HOIL-1L (RBCK1), and SHARPIN [38]. HOIP contains the catalytic RBR (RING-IBR-RING) domain and the unique LDD (Linear ubiquitin chain determining Domain) that positions the acceptor ubiquitin for M1-linkage [60] [38]. HOIL-1L and SHARPIN stabilize the complex and allosterically enhance HOIP's catalytic activity [38]. LUBAC is recruited to signaling complexes, such as the TNFR1 complex I and the LTβR signaling complex, where it linearly ubiquitinates substrates like NEMO (IKKγ) and RIPK1 [57] [56]. These chains then serve as a platform for the recruitment and activation of the IKK complex, leading to NF-κB activation and the transcription of pro-inflammatory and pro-survival genes [57] [56] [60].
OTULIN (The Eraser): OTULIN is a deubiquitinase (DUB) that exclusively hydrolyzes linear ubiquitin chains [56] [4]. It interacts with the PUB (Peptide using N-terminal ubiquitin) domain of HOIP, placing it in close proximity to LUBAC to tightly regulate its activity and prevent aberrant signaling [38] [4]. OTULIN's role extends beyond mere chain disassembly; recent paradigm-shifting research has revealed that OTULIN also functions as a master regulator of gene expression and RNA metabolism [58] [59] [61]. OTULIN deficiency in neurons leads to the complete disappearance of tau mRNA, indicating a non-canonical role in controlling mRNA stability and transcription that is independent of its deubiquitinase activity [58] [59] [61].
Readers: The functional outcome of linear ubiquitination is mediated by proteins that specifically recognize this chain type. The most prominent reader is NEMO, which binds linear chains via its UBAN (Ubiquitin binding in ABIN and NEMO) domain, thereby activating the IKK complex [56].
The following diagram illustrates the core linear ubiquitination machinery and its primary role in the NF-κB signaling pathway.
Therapeutic Rationale for Agonists and Inhibitors
The central role of LUBAC and OTULIN in human pathology makes them compelling drug targets.
LUBAC as a Target for Agonists and Inhibitors: The therapeutic strategy for LUBAC modulation is context-dependent. In certain cancers, such as hepatocellular carcinoma (HCC) with high LTβR expression, LUBAC activity promotes a tumor-promoting, inflammatory secretome. In these contexts, LUBAC inhibitors may be beneficial [57]. Conversely, in specific immunodeficiencies or scenarios requiring enhanced immune activation, LUBAC agonists could be therapeutic. However, the development of true agonists that directly enhance LUBAC's E3 ligase activity remains a significant challenge.
OTULIN as a Target for Inhibitors: Inhibiting OTULIN is a promising strategy for several reasons. First, it would lead to a net increase in linear ubiquitination, potentially augmenting immune signaling in certain contexts. Second, and more notably, its newly discovered role in tau expression opens a transformative avenue for treating Alzheimer's disease and other tauopathies. Partial inhibition of OTULIN with a small molecule (UC495) was shown to reduce pathological phosphorylated tau without eliminating total tau, suggesting a viable therapeutic window [58] [59] [61]. Furthermore, OTULIN has been shown to protect against acetaminophen (APAP)-induced liver injury by modulating ferroptosis, indicating potential applications in hepatoprotection [62].
Quantitative Data on Known Modulators
The field has developed several pharmacological tools, primarily focused on LUBAC inhibition. The quantitative data for key compounds are summarized in the table below.
Table 1: Quantitative Profiling of Characterized LUBAC Inhibitors
| Compound Name | Chemical Class | Molecular Target | Reported IC₅₀ / Efficacy | Cellular & Phenotypic Effects |
|---|---|---|---|---|
| HOIPIN-1 [60] | α,β-unsaturated carbonyl | Covalently modifies HOIP's active site Cys885 | Suppresses IL-1β-induced linear ubiquitination and NF-κB activation in A549 cells. | Reduces pro-inflammatory gene expression; does not affect MAPK pathway. |
| HOIPIN-8 [60] | Derivative of HOIPIN-1 | Covalently modifies HOIP's active site Cys885 | More potent than HOIPIN-1 in suppressing NF-κB activation. | Induces apoptosis in ABC-DLBCL cells; alleviates psoriasis in mouse models. |
| UC495 [58] [59] [61] | Novel small-molecule (structure not fully disclosed) | OTULIN deubiquitinase inhibitor | Reduces phosphorylated tau levels in patient-derived Alzheimer's neurons. | Partial inhibition reduces pathological tau without neuronal toxicity; complete knockout ablates tau production. |
| BAY11-7082 [60] | Vinyl sulfone | Non-specific, thiol-reactive | Not a specific LUBAC inhibitor at non-toxic doses. | General NF-κB inhibition with high cytotoxicity; not recommended for LUBAC-specific studies. |
| Gliotoxin [60] | Mycotoxin | Non-specific, thiol-reactive | Not a specific LUBAC inhibitor at non-toxic doses. | General NF-κB and JNK inhibition; high cytotoxicity. |
Experimental Protocols for Screening and Validation
This section outlines detailed methodologies for key experiments in the discovery and validation of LUBAC and OTULIN modulators.
In Vitro Biochemical Assays
LUBAC Linear Ubiquitination Assay
Purpose: To screen for and characterize direct inhibitors or agonists of LUBAC's E3 ligase activity. Protocol:
- Reagent Preparation: Purify recombinant trimeric LUBAC complex (HOIP/HOIL-1L/SHARPIN). Prepare E1 (UBA1), E2 (UBE2L3), ubiquitin, and an energy-regenerating system (ATP, Mg²⁺) in reaction buffer [60] [38].
- Reaction Setup: In a 50 µL reaction, mix LUBAC (50 nM), E1 (100 nM), E2 (1 µM), ubiquitin (10 µM), and the test compound. Pre-incubate the enzyme-compound mixture for 15 minutes on ice before initiating the reaction with ATP.
- Incubation: Incubate at 30°C for 60-90 minutes.
- Termination & Analysis: Quench the reaction with SDS-PAGE loading buffer. Analyze the formation of high-molecular-weight linear ubiquitin chains by:
- Western Blotting: Using a linkage-specific antibody for linear (M1) ubiquitin.
- Gel Staining: Visualize total ubiquitin polymers using Coomassie or silver stain.
OTULIN Deubiquitinase Assay
Purpose: To screen for and characterize inhibitors of OTULIN's enzymatic activity. Protocol:
- Substrate Preparation: Generate linear polyubiquitin chains using purified LUBAC in vitro or purchase commercially available substrates.
- Reaction Setup: In a 50 µL reaction, mix the linear ubiquitin substrate (1 µM) with purified OTULIN (50 nM) and the test compound in DUB assay buffer.
- Incubation: Incubate at 37°C for 30-60 minutes.
- Termination & Analysis: Quench with SDS-PAGE loading buffer. Analyze the disappearance of linear ubiquitin chains and appearance of free ubiquitin by:
- Western Blotting: Using anti-linear ubiquitin and anti-free ubiquitin antibodies.
- Fluorescence-Based Assay: Use ubiquitin chains with a C-terminal fluorescent tag (e.g., Rhodamine); cleavage by OTULIN results in a quantifiable fluorescence increase [56].
Cell-Based Functional Assays
NF-κB Pathway Activation/Inhibition Assay
Purpose: To assess the functional cellular consequences of LUBAC or OTULIN modulation on canonical NF-κB signaling. Protocol:
- Cell Culture & Treatment: Use relevant cell lines (e.g., A549, HeLa, Jurkat T-cells). Pre-treat cells with the compound for a predetermined time (e.g., 2-4 hours).
- Stimulation: Activate the pathway with a relevant stimulus (e.g., TNF-α, IL-1β, LIGHT) for 5-30 minutes [57] [60].
- Cell Lysis & Western Blotting: Lyse cells and analyze key signaling events by Western blot.
- Primary Antibodies: Phospho-IκBα, total IκBα, Phospho-IKKα/β, Phospho-p65, and linear ubiquitin.
- Loading Controls: β-actin or GAPDH.
Monitoring Tau Expression in Neuronal Models
Purpose: To validate OTULIN inhibitors in the context of Alzheimer's disease and tauopathies. Protocol:
- Cell Models: Use human neuroblastoma cell lines (e.g., SH-SY5Y) or induced pluripotent stem cell (iPSC)-derived neurons from Alzheimer's patients and healthy controls [58] [59] [61].
- Genetic or Pharmacological Manipulation:
- Knockout: Use CRISPR/Cas9 to generate OTULIN-deficient cell lines.
- Inhibition: Treat cells with the OTULIN inhibitor UC495.
- Downstream Analysis:
- Western Blotting: Quantify total tau and phosphorylated tau levels.
- qRT-PCR: Measure MAPT (tau) mRNA levels to confirm transcriptional/RNA stability regulation.
- RNA-Sequencing: Perform bulk RNA-Seq to comprehensively analyze global changes in gene expression and RNA metabolism, as performed in the seminal study [61].
The following diagram outlines a typical workflow for screening and validating modulators.
The Scientist's Toolkit: Key Research Reagents
Successful research in this field relies on a suite of specific reagents and tools, as cataloged below.
Table 2: Essential Research Reagents for LUBAC and OTULIN Studies
| Reagent Category | Specific Example | Function and Application in Research |
|---|---|---|
| Linkage-Specific Antibodies | Anti-linear (M1) ubiquitin antibody | Critical for detecting endogenous linear ubiquitination by Western blotting and immunofluorescence; used to monitor LUBAC activity and OTULIN inhibition [60]. |
| Validated Cell Lines | HOIP-deficient Jurkat T-cells | Isogenic control cell lines to confirm the on-target effects of LUBAC-targeting compounds and rule off-target effects [60]. |
| OTULIN KO lines (e.g., SH-SY5Y) | Used to elucidate the non-canonical role of OTULIN in RNA metabolism and tau regulation [58] [61]. | |
| Recombinant Proteins | Purified trimeric LUBAC complex | Essential for in vitro ubiquitination assays and biochemical characterization of inhibitors [60] [38]. |
| Recombinant OTULIN (full-length and OTU domain) | Required for DUB activity assays and structural studies (e.g., crystallography) to guide inhibitor design [56] [4]. | |
| Specialized Substrates | In vitro generated linear polyubiquitin chains | The specific substrate for OTULIN DUB activity assays [56]. |
| Pathway Reporter Systems | NF-κB luciferase reporter gene | Allows for quantitative, high-throughput screening of compounds that modulate the LUBAC-dependent NF-κB pathway [60]. |
| Patient-Derived Models | iPSC-derived neurons from Alzheimer's patients | Provide a physiologically relevant human model for testing the efficacy of OTULIN inhibitors in reducing pathological tau [59] [61]. |
The drug discovery landscape for LUBAC and OTULIN modulators is rapidly evolving, moving from basic tool compounds to candidates with therapeutic potential. The discovery of HOIPINs validates LUBAC as a druggable target for inflammatory diseases and cancer, while the identification of UC495 and the elucidation of OTULIN's role in tau biology open a new frontier for treating neurodegenerative diseases. Future efforts should focus on developing more potent and specific compounds, including the non-covalent and allosteric inhibitors for LUBAC, and agonists that could fine-tune its activity. For OTULIN, the major challenge and opportunity lie in exploiting its dual functions—modulating its deubiquitinase activity to control immune signaling versus targeting its non-canonical role in RNA metabolism to combat tauopathies. As our understanding of the complex biology of linear ubiquitination deepens, so too will the sophistication and promise of its associated pharmacopeia.
Resolving Experimental Challenges in Linear Ubiquitin Research and Pathway Modulation
M1-linked, or linear, ubiquitination is a crucial post-translational modification characterized by a head-to-tail linkage of ubiquitin molecules via the N-terminal methionine (M1) of the acceptor ubiquitin and the C-terminal glycine of the donor ubiquitin [38]. Unlike other ubiquitin linkages that form isopeptide bonds, linear ubiquitination creates a unique peptide bond, resulting in distinct structural and functional properties [38]. The linear ubiquitin chain assembly complex (LUBAC), comprising HOIP, HOIL-1L, and SHARPIN, serves as the sole E3 ligase capable of generating these chains, with HOIP containing the catalytically essential linear ubiquitin chain determining domain (LDD) [38] [4]. This modification plays an indispensable role in regulating immune signaling pathways, particularly NF-κB activation, with demonstrated importance in inflammation, cell death regulation, and immunity [31] [63] [4].
The fundamental challenge in studying M1-linked ubiquitination stems from its intrinsically transient nature and low stoichiometry within cells. M1-ubiquitin chains are generated in a highly spatially and temporally regulated manner, often as rapid response elements to signaling cues [38]. This transient existence is compounded by the activity of dedicated deubiquitinases (DUBs), specifically OTULIN and CYLD, which efficiently disassemble these chains [38] [4]. Furthermore, the low abundance of M1-chain modifications relative to total cellular protein creates significant detection hurdles, as the window for capturing these events is narrow before DUB-mediated reversal or substrate degradation occurs [64]. This technical barrier has limited our understanding of the full scope of M1-linked ubiquitination in physiology and disease, necessitating robust methodological approaches for stabilization and detection.
Understanding the M1-Linked Ubiquitination Machinery
Writers, Erasers, and Readers of the Linear Ubiquitin Code
The linear ubiquitin code is dynamically regulated by a precise enzymatic machinery: writers (LUBAC) that install the modification, erasers (DUBs) that remove it, and readers that interpret the signal to produce functional outcomes [38].
Writer Complex - LUBAC: The LUBAC complex exhibits a sophisticated architecture where HOIP serves as the catalytic core, while HOIL-1L and SHARPIN play crucial structural and regulatory roles. The interaction between the ubiquitin-associated (UBA) domain of HOIP and ubiquitin-like (UBL) domains of HOIL-1L and SHARPIN relieves HOIP's autoinhibition and stabilizes the active complex [38]. Recent research has demonstrated that STING activation recruits HOIP to LC3B-associated Golgi membranes, where it synthesizes M1-ubiquitin chains to stimulate both NF-κB and IRF3 signaling pathways, expanding the known locales of linear ubiquitination beyond traditional signaling complexes [31].
Erasers - OTULIN and CYLD: These DUBs provide the counter-regulatory mechanism that ensures signaling transientness. OTULIN exhibits remarkable specificity for linear chains through its ovarian tumor (OTU) domain, which contains conserved residues (Cys129, His339, Asn341) that confer selective recognition of M1-linkages [4]. The interaction between OTULIN's PUB-interacting motif (PIM) and the PUB domain of HOIP facilitates regulated deubiquitination, with phosphorylation at Tyr56 negatively modulating this interaction [4].
Readers - Ubiquitin Binding Domains: Proteins equipped with specific ubiquitin-binding domains (UBDs), such as NEMO (NF-κB essential modulator), function as readers that translate the M1-ubiquitin signal into downstream cellular responses. Recent findings indicate that M1-ubiquitin chains induce phase separation of NEMO, promoting the formation of biomolecular condensates that facilitate efficient NF-κB pathway activation [65].
Table 1: Core Components of the Linear Ubiquitination Machinery
| Component | Elements | Primary Function | Key Features |
|---|---|---|---|
| Writer | LUBAC Complex: HOIP, HOIL-1L, SHARPIN | M1-linked ubiquitin chain assembly | HOIP contains catalytic RBR and LDD domains; Sole E3 for linear chains |
| Erasers | OTULIN, CYLD | Removal of M1-linked ubiquitin chains | OTULIN highly specific for M1-linkage; CYLD has broader specificity |
| Readers | NEMO, ABIN proteins, OPTN | Signal transduction via UBDs | NZF and UBAN domains recognize M1 topology; Induce phase separation |
Biological Significance in Immune Signaling and Disease
Linear ubiquitination serves as a critical regulatory mechanism across multiple immune signaling pathways. In NF-κB signaling, M1-ubiquitin chains formed by LUBAC enable the recruitment and activation of the IKK complex through NEMO binding, leading to IκBα phosphorylation and degradation, and subsequent nuclear translocation of NF-κB transcription factors [65] [4]. Beyond this canonical pathway, emerging roles for linear ubiquitination include regulation of cell death pathways (apoptosis, necroptosis), autophagy, and responses to intracellular pathogens [63] [4].
Recent findings have revealed unexpected functions in organelle-specific signaling, including evidence that linear ubiquitination at damaged lysosomes induces local NF-κB activation and controls cell survival decisions [63]. Similarly, mitochondrial outer membrane integrity has been shown to regulate a ubiquitin-dependent and NF-κB-mediated inflammatory response, potentially involving M1-chain modifications [63]. The functional importance of this system is underscored by human disease manifestations: mutations in LUBAC components cause severe autoinflammatory diseases, while OTULIN deficiency leads to ORAS (OTULIN-related autoinflammatory syndrome) [4].
Methodological Framework for Stabilizing M1-Linked Ubiquitin Chains
Inhibition of Deubiquitinating Enzymes (DUBs)
Strategic pharmacological inhibition of the M1-specific DUBs OTULIN and CYLD represents the most direct approach to stabilize endogenous linear ubiquitin chains. The effectiveness of DUB inhibition is highly dependent on proper inhibitor selection and application timing.
Cell Permeable Broad-Spectrum DUB Inhibitors: PR-619 serves as a valuable first-line tool for initial stabilization experiments. This cell-permeable inhibitor targets cysteine-dependent DUBs, including OTULIN and CYLD, with an effective concentration range of 10-50 μM for cellular treatments. Treatment duration should typically range from 2-6 hours before sample collection, as longer exposures may induce compensatory mechanisms or non-specific toxicity [64]. PR-619 is particularly useful when combined with proteasome inhibitors like MG-132 or bortezomib to prevent degradation of ubiquitinated substrates.
Specific DUB Inhibition Strategies: While no highly specific commercially available inhibitors exist exclusively for OTULIN, targeting the related DUB CYLD can be achieved using compounds such thanatin or specific siRNA knockdown approaches. For the most comprehensive stabilization, combined targeting of both OTULIN and CYLD through genetic approaches (siRNA/shRNA) provides maximal M1-chain accumulation, though this may potentially disrupt normal signaling feedback mechanisms [4].
Critical Considerations for DUB Inhibition: Researchers must include appropriate DUB inhibitor controls, including vehicle-treated samples and specificity controls where possible. Timing of inhibitor application is crucial—addition too early may alter baseline signaling, while addition too late may miss transient M1-chain formation. Optimal results are obtained when inhibitors are applied simultaneously with or immediately following pathway stimulation.
Table 2: DUB Inhibitors for M1-Chain Stabilization
| Inhibitor | Specificity | Working Concentration | Advantages | Limitations |
|---|---|---|---|---|
| PR-619 | Broad-spectrum cysteine DUBs | 10-50 μM (cellular) | Cell-permeable; Effective for OTULIN/CYLD | Limited specificity; Potential off-target effects |
| siRNA/shRNA | OTULIN or CYLD | Gene-specific knockdown | High specificity; Suitable for chronic studies | Requires 48-72 hr transfection; Compensation possible |
| Thanatin | CYLD-preferred | Literature-dependent | More specific than broad inhibitors | Limited commercial availability; Not OTULIN-specific |
Genetic and Molecular Manipulation of LUBAC Activity
Complementary to DUB inhibition, enhancing LUBAC-mediated M1-chain assembly through genetic and molecular approaches provides an alternative stabilization strategy.
LUBAC Overexpression: Transient or stable expression of LUBAC components, particularly wild-type HOIP, enhances basal M1-chain formation. The catalytically active RBR-LDD fragment of HOIP (lacking the autoinhibitory N-terminal domain) demonstrates significantly increased activity compared to full-length HOIP [38]. Co-expression of all three LUBAC components (HOIP, HOIL-1L, and SHARPIN) typically yields optimal results, as the complex requires all subunits for full activity and stability [38].
Pathway-Specific Stimulation: Combining LUBAC overexpression with pathway-specific agonists maximizes M1-chain detection. For NF-κB pathways, stimulation with TNF-α (10-100 ng/mL for 5-30 minutes) or IL-1β (10-50 ng/mL for 5-30 minutes) robustly induces linear ubiquitination [65]. Recent research indicates that STING activation by cyclic dinucleotides also induces HOIP-mediated M1-chain synthesis on Golgi membranes, providing an additional pathway for stimulation [31].
Constitutive Active Mutants: Engineered LUBAC variants, such as N-terminally truncated HOIP (ΔN-HOIP) that lacks autoinhibitory domains, can produce sustained M1-chain formation without extracellular stimulation [38]. However, researchers should exercise caution as constitutive signaling may alter physiological feedback mechanisms and potentially induce cytotoxicity.
Sample Preparation and Lysis Conditions for M1-Chain Preservation
The successful stabilization and detection of M1-linked ubiquitin chains is critically dependent on appropriate sample preparation and lysis conditions that preserve these labile modifications.
Lysis Buffer Composition: The standard practice of including protease inhibitors in lysis buffers must be expanded to include dedicated DUB inhibitors at sufficient concentrations. Recommended lysis buffers should contain:
- Metalloprotease DUB Inhibitors: EDTA/EGTA (1-5 mM) to target metal-dependent DUBs
- Cysteine Protease DUB Inhibitors: N-ethylmaleimide (NEM; 1-10 mM), iodoacetamide (5-20 mM), or 2-chloroacetamide (5-15 mM) to inhibit cysteine-dependent DUBs like OTULIN and CYLD
- Broad-Spectrum DUB Inhibitors: PR-619 (10-50 μM) can be included in lysis buffers for additional protection [64]
Lysis Conditions and Handling: Maintaining samples at low temperatures (0-4°C) throughout the lysis procedure is essential to minimize DUB activity. Rapid processing and immediate freezing of samples in liquid nitrogen followed by storage at -80°C helps preserve ubiquitination states. For tissue samples, particularly skeletal muscle, rapid dissection and freezing within minutes of collection is critical [64] [66].
Special Considerations for Subcellular Fractionation: As M1-chains form in specific subcellular compartments (e.g., Golgi membranes, mitochondrial-associated membranes), fractionation protocols may enhance detection sensitivity [31] [63]. These protocols require the same DUB inhibition strategies applied to homogenization and fractionation buffers to prevent chain disassembly during processing.
Advanced Detection and Quantification Methods for M1-Chains
Enrichment Strategies for Low-Abundance M1-Chains
Effective detection of stabilized M1-chains requires specialized enrichment strategies to overcome their low stoichiometry relative to total cellular protein.
Tandem Ubiquitin Binding Entities (TUBEs): These engineered reagents comprise multiple ubiquitin-binding domains (UBDs) arranged in tandem, creating avidity effects that significantly enhance affinity for polyubiquitin chains. TUBEs based on domains with preference for M1-linkages (e.g., UBAN domains from NEMO) offer improved selectivity for linear chains. The MultiDsk system, incorporating multiple UBDs from the yeast protein Dsk2, demonstrates particularly effective enrichment of diverse ubiquitin chain types, including M1-linkages [64].
Linkage-Specific Antibodies: Antibodies specifically recognizing M1-linked ubiquitin chains provide the most direct enrichment method. Commercial M1-specific antibodies (e.g., from Millipore, Cell Signaling Technology) enable immunoprecipitation of linear ubiquitinated proteins, though careful validation is required to confirm specificity, particularly in distinguishing M1-linkages from other chain types. These antibodies can be used in conjunction with TUBEs for sequential enrichment to maximize purification of M1-chains.
Modified Ubiquitin Traps: Expression of ubiquitin-binding proteins fused to solid support tags (e.g., GST-UBAN domains, His-NZF arrays) enables affinity purification of M1-chains from cell lysates. Co-expression of these traps with LUBAC components and stimulus application can enhance capture efficiency, particularly for transient signaling events.
Detection and Analytical Techniques
Following enrichment, multiple analytical approaches enable detection and quantification of M1-linked ubiquitin chains.
Immunoblotting with Linkage-Specific Antibodies: Western blotting remains the most accessible detection method, with M1-specific antibodies allowing visualization of free M1-chains and M1-modified substrates. Key considerations include:
- Sample denaturation at 95°C for 5-10 minutes in SDS-containing buffer to disrupt non-covalent interactions
- Inclusion of positive controls (LUBAC overexpression) and negative controls (OTULIN overexpression or LUBAC knockdown)
- Comparison with total ubiquitin levels to assess relative abundance
Mass Spectrometry-Based Ubiquitylomics: For comprehensive mapping of M1-ubiquitination sites, mass spectrometry approaches following diGly remnant immunoprecipitation provide the most powerful solution [64]. This methodology involves:
- Expression of epitope-tagged ubiquitin or use of endogenous ubiquitin detection
- Trypsin digestion generating diGly lysine remnants (K-ε-GG) from ubiquitinated sites
- Immunoaffinity enrichment of diGly-modified peptides using specific antibodies
- LC-MS/MS analysis for identification and quantification Specialized computational tools then help distinguish M1-linkage signatures from other ubiquitin chain types based on unique peptide patterns.
Advanced Microscopy and Cellular Visualization: For spatial analysis of M1-chain formation, immunofluorescence staining of fixed cells using M1-linkage specific antibodies enables subcellular localization. Recent super-resolution techniques have revealed that M1-ubiquitin chains induce phase separation of NEMO into biomolecular condensates, providing insights into the mechanism of signal amplification [65]. Proximity ligation assays (PLA) can visualize specific protein interactions mediated by M1-chains with subcellular resolution.
Table 3: Key Research Reagents for M1-Chain Studies
| Reagent Category | Specific Examples | Application | Considerations |
|---|---|---|---|
| DUB Inhibitors | PR-619, N-Ethylmaleimide (NEM), Iodoacetamide | Stabilization of endogenous M1-chains | Varying specificity; Potential off-target effects |
| LUBAC Expression Constructs | Wild-type HOIP, ΔN-HOIP (constitutive active), Full LUBAC complex | Enhanced M1-chain generation | May overwhelm endogenous regulation |
| M1-Specific Antibodies | Anti-linear ubiquitin (Millipore MABS199), Anti-K48-linkage, Anti-K63-linkage | Detection, quantification, enrichment | Must validate specificity for intended applications |
| Ubiquitin Binders | TUBEs, MultiDsk, OtUBD, UBAN-domain fusions | Enrichment of ubiquitinated proteins | Differing linkage preferences and affinities |
| Activity Reporters | NEMO-UBAN fusions, Luciferase-NF-κB reporters | Functional assessment of M1-signaling | Measures downstream output rather than direct M1-chains |
Applications in Disease Research and Therapeutic Development
The methodological advances in stabilizing and detecting M1-linked ubiquitin chains have enabled deeper investigation into their roles in human disease pathophysiology and created opportunities for therapeutic intervention.
In autoimmune and inflammatory diseases, aberrant linear ubiquitination contributes to dysregulated immune responses. Rheumatoid arthritis research has revealed that exosomal circ-CBLB promotes M1 macrophage polarization through the TLR3/TRAF3 signaling axis, suggesting potential points for intervention in the ubiquitination cascade [67]. Similarly, in OTULIN-related autoinflammatory syndrome (ORAS), mutations in the M1-specific deubiquitinase OTULIN lead to uncontrolled linear ubiquitination and severe inflammation, highlighting the critical balance required in this pathway [4].
Cancer biology has revealed connections between linear ubiquitination and tumor progression. Gastric cancer studies identified an M1 macrophage-related signature that predicts prognosis and therapeutic response, with fibronectin 1 (FN1) emerging as a hub gene with oncogenic functions [68]. The role of linear ubiquitination in regulating multiple cell death modalities, including apoptosis, necroptosis, and ferroptosis, positions the LUBAC complex as a potential target for cancer therapy [4].
In infectious disease and immunodeficiency, recent work has demonstrated that linear ubiquitination of cytosolic Salmonella Typhimurium activates NF-κB and restricts bacterial proliferation, revealing an antimicrobial function for this modification [63]. Additionally, research in people living with HIV has uncovered mitochondrial respiratory chain deficiencies associated with inflammation and aberrant muscle regeneration, suggesting potential connections to ubiquitin-mediated signaling in wasting conditions [66].
Emerging therapeutic approaches targeting the linear ubiquitination pathway include:
- LUBAC Inhibitors: Development of small molecule inhibitors targeting HOIP's catalytic activity, such as HOIPIN-1 and derivatives, show promise in preclinical models of inflammation and cancer
- Protein-Targeted Degradation: PROTAC molecules designed to specifically degrade hyperactive LUBAC components in autoimmune conditions
- Stabilization Strategies: Compounds that enhance linear ubiquitination in immunodeficiency disorders or specific infectious diseases
These therapeutic avenues highlight the translational potential of understanding and manipulating the linear ubiquitination pathway, with methodologies for stabilizing M1-chains playing a crucial role in both basic research and drug development.
The stabilization of transient and low-abundance M1-linked ubiquitin chains represents a critical methodological challenge that must be overcome to fully elucidate the biological functions of this unique post-translational modification. The integrated approach combining pharmacological inhibition of DUBs, optimized sample preparation, and sensitive enrichment strategies provides a robust framework for researchers to capture these elusive signaling events. As these methodologies continue to evolve, several emerging areas promise to further advance the field.
Future developments will likely include the creation of more specific OTULIN inhibitors that avoid off-target effects on other cysteine DUBs, the engineering of next-generation TUBEs with enhanced specificity for M1-linkages, and the implementation of real-time biosensors for monitoring M1-chain dynamics in living cells. Additionally, the application of single-cell ubiquitylomics approaches may reveal cell-to-cell heterogeneity in linear ubiquitination that is masked in bulk analyses.
The continued refinement of these stabilization and detection methodologies will undoubtedly expand our understanding of the diverse physiological processes regulated by linear ubiquitination and accelerate the development of therapeutics targeting this pathway in human disease. As research progresses, the insights gained from studying M1-linked ubiquitin chains will continue to illuminate fundamental principles of cellular signaling and immune regulation.
Ubiquitination is a sophisticated post-translational modification that regulates vast cellular processes, from protein degradation to immune signaling. Its versatility stems from the capacity to form polyubiquitin chains through different linkages between ubiquitin monomers, creating a complex "ubiquitin code" that determines specific biological outcomes [38]. Among these linkages, M1-linked (linear) and K63-linked ubiquitin chains have emerged as crucial regulators of innate and adaptive immune responses. Furthermore, branched ubiquitin chains, which contain more than one linkage type, add another layer of complexity to this regulatory system [69]. This technical guide provides an in-depth analysis of the structural, functional, and methodological distinctions between these ubiquitin chain types, with particular emphasis on M1-linear ubiquitination within immune signaling research.
Structural and Functional Distinctions Between Chain Types
The fundamental differences between M1-linked, K63-linked, and branched ubiquitin chains define their unique roles in cellular physiology, particularly in the regulation of immune signaling pathways.
M1-Linked Linear Ubiquitin Chains
M1-linked (linear) ubiquitin chains are characterized by a unique peptide bond formed between the C-terminal glycine of the donor ubiquitin and the α-amino group of the N-terminal methionine (M1) of the acceptor ubiquitin [38]. This linear architecture is exclusively assembled by the Linear Ubiquitin Chain Assembly Complex (LUBAC), a multi-subunit E3 ubiquitin ligase complex comprising HOIP, HOIL-1, and SHARPIN [70] [38]. HOIP serves as the catalytically active component, with its unique C-terminal Linear Ubiquitin Chain Determining Domain (LDD) positioning the N-terminus of the target ubiquitin for M1-linkage formation [38]. Functionally, linear ubiquitin chains are crucial modulators of inflammatory signaling and cell death pathways, primarily through their role in regulating TNF and NF-κB signaling cascades [70].
K63-Linked Ubiquitin Chains
In contrast, K63-linked ubiquitin chains are formed through a conventional isopeptide bond between the C-terminal glycine of the donor ubiquitin and the ε-amino group of lysine 63 (K63) on the acceptor ubiquitin. These chains are synthesized by various E2/E3 enzyme pairs, with the Ubc13-Uev1a E2 complex in conjunction with TRAF6 E3 ligase being particularly important in immune signaling contexts [71]. Structurally, K63-linked chains adopt a more open, extended conformation compared to the compact structures of K48-linked chains. Their primary function in immune signaling involves mediating protein-protein interactions and facilitating the assembly of signaling complexes rather than targeting proteins for degradation [71].
Branched Ubiquitin Chains
Branched ubiquitin chains represent a more complex category, characterized by ubiquitin monomers that are simultaneously modified on at least two different acceptor sites [69]. These heterotypic chains can incorporate various linkage combinations, with K48/K63, K11/K48, and K29/K48 branched architectures being among the best characterized. The synthesis of branched chains often involves collaboration between pairs of E3 ligases with distinct linkage specificities or single E3s that can recruit E2s with different linkage preferences [69]. For instance, during NF-κB signaling, TRAF6 and HUWE1 collaborate to produce branched K48/K63 chains, while the APC/C cooperates with UBE2C and UBE2S E2 enzymes to form branched K11/K48 chains during mitosis [69]. Functionally, branched chains can act as particularly potent degradation signals or facilitate the conversion of non-degradative signals to degradative marks [69].
Table 1: Key Characteristics of Ubiquitin Chain Types
| Characteristic | M1-Linked (Linear) | K63-Linked | Branched (K48/K63) |
|---|---|---|---|
| Bond Type | Peptide bond | Isopeptide bond | Multiple isopeptide bonds (and potentially peptide bonds) |
| Forming Enzyme | LUBAC (HOIP/HOIL-1/Sharpin) | Various E2/E3 pairs (e.g., Ubc13-TRAF6) | Collaborative E3 pairs (e.g., TRAF6/HUWE1) or specific E3s (e.g., UBR5) |
| 3D Structure | Extended, open conformation | Extended, open conformation | Complex, compact structures |
| Primary Immune Function | Regulation of TNF and NF-κB signaling; inflammatory and cell death signaling | Scaffold for signaling complex assembly in IL-1R/TLR pathways | Potent degradative signal; regulation of signaling amplitude and duration |
| Key Binding Proteins | NEMO (high affinity) | TAB2/TAB3 (TAK1 complex) | Multiple proteasomal receptors |
| Chain Disassembly | Otulin DUB | AMSH-LP DUB | Multiple DUBs often required |
Diagram 1: Structural and enzymatic differences between ubiquitin chain types.
Specific Roles in Immune Signaling Pathways
M1-Linear Ubiquitination in NF-κB Activation
The role of M1-linear ubiquitination in immune signaling is particularly well-defined in the context of TNF receptor and IL-1R/TLR signaling pathways. LUBAC-generated linear ubiquitin chains are recruited to signaling complexes where they directly bind to NEMO (NF-κB Essential Modifier), a regulatory component of the IκB kinase (IKK) complex [70] [71]. This interaction occurs with remarkably high affinity—approximately 100-fold greater than NEMO's affinity for K63-linked chains [71]. The binding of linear chains to NEMo facilitates the conformational activation of the IKK complex, leading to phosphorylation of IκB and subsequent activation of NF-κB transcription factors. Beyond NF-κB activation, linear ubiquitination also plays a critical role in regulating cell death signaling pathways, including both apoptosis and necroptosis, thereby fine-tuning the balance between inflammatory gene activation and cell survival decisions [70].
K63-Ubiquitination in Early Signaling Events
K63-linked ubiquitin chains function earlier in immune signaling cascades, particularly in the MyD88-dependent signaling network activated by IL-1R and TLR engagement [71]. Following receptor activation, the adaptor proteins MyD88, IRAK4, and IRAK1 form an oligomeric structure termed the Myddosome. IRAK1 then undergoes K63-linked ubiquitylation, potentially through the action of TRAF6 and the Ubc13-Uev1a E2 complex [71]. These K63-linked chains serve as platforms for the recruitment of the TAK1 kinase complex through binding to its regulatory subunits TAB2 and TAB3. The TAK1 complex, once recruited and activated, phosphorylates and activates the IKK complex as well as MAP kinase kinases, thereby initiating both NF-κB and MAPK signaling arms [71].
Hybrid and Branched Chains in Signal Integration
Recent research has revealed that hybrid and branched ubiquitin chains play sophisticated roles in immune signal integration and amplification. A key discovery is that nearly all M1-pUb chains formed in response to IL-1 or TLR agonists are covalently attached to K63-pUb chains, either directly as K63/M1-pUb hybrids or indirectly by attachment to the same protein [71]. This hierarchical assembly occurs sequentially: IRAK1 is modified first by K63-pUb chains, to which M1-pUb linkages are subsequently added by LUBAC [71]. The formation of K63-pUb chains is actually a prerequisite for the subsequent formation of M1-pUb chains, with HOIP specifically recognizing and binding to K63-pUb chains to initiate linear chain assembly [71]. These hybrid chains function as sophisticated scaffolds that colocalize the TAK1 and IKK complexes, facilitating the TAK1-catalyzed activation of IKK subunits and ensuring rapid, coordinated signaling in response to immune stimuli [71].
Diagram 2: Sequential ubiquitination in IL-1R/TLR signaling leading to NF-κB activation.
Methodologies for Detection and Distinction
Linkage-Specific Reagents and Detection Methods
The discrimination between different ubiquitin chain types requires specialized reagents and methodological approaches that exploit the unique structural features and binding preferences of each linkage type.
Table 2: Detection Methodologies for Ubiquitin Chain Types
| Methodology | M1-Linear Specificity | K63-Linkage Specificity | Branched Chain Detection |
|---|---|---|---|
| Linkage-Specific Antibodies | Monoclonal antibodies specific for linear ubiquitin (e.g., M1-specific) | K63-linkage specific antibodies | Limited availability; often requires combination of linkage-specific antibodies |
| Ubiquitin-Binding Domains (UBDs) | NEMO NZF domain (high affinity) | TAB2/TAB3 NZF domains | Tandem UBD arrangements with different specificities |
| Deubiquitinase (DUB) Profiling | Otulin (high specificity) | AMSH-LP (high specificity) | Sequential DUB treatments required |
| Mass Spectrometry | Identification of M1-linked peptides after trypsin digestion (generates unique signature) | Identification of K63-linked peptides | Advanced middle-down or intact protein MS with electron-transfer dissociation |
| Biochemical Pull-Down | Halo-NEMO beads | Tandem Ubiquitin Binding Entities (TUBEs) | Modified TUBE approaches with different linkage preferences |
Experimental Workflow for Distinguishing Chain Types
The following detailed experimental protocol, adapted from Emmerich et al. (2013), allows for the specific detection and distinction of M1-linear and K63-linked ubiquitin chains in immune signaling contexts [71]:
Cell Stimulation and Lysis:
- Culture IL-1R-expressing HEK293 cells in appropriate medium.
- Stimulate cells with IL-1β (10-100 ng/mL) for varying time points (0-30 minutes).
- Lyse cells in ice-cold lysis buffer (e.g., RIPA buffer) supplemented with protease inhibitors (including 10 μM MG132 to prevent proteasomal degradation) and 20 mM N-ethylmaleimide to inhibit endogenous DUBs.
Ubiquitin Chain Capture:
- Incubate clarified cell lysates with Halo-NEMO fusion protein immobilized on HaloLink resin.
- As critical controls, include parallel samples using:
- Halo-NEMO[D311N] mutant (ubiquitin-binding deficient) to confirm specificity
- Halo-tandem ubiquitin-binding entities (TUBEs) for broader ubiquitin chain capture
- Perform binding for 2-4 hours at 4°C with gentle rotation.
DUB-Based Linkage Specificity Analysis:
- Divide captured ubiquitin chains into three aliquots:
- Untreated control: Incubate with reaction buffer only
- Otulin treatment: Incubate with 1 μM Otulin (M1-specific DUB) for 1 hour at 37°C
- AMSH-LP treatment: Incubate with 1 μM AMSH-LP (K63-specific DUB) for 1 hour at 37°C
- Stop reactions by adding SDS-PAGE loading buffer with DTT.
Immunoblot Analysis:
- Resolve samples by SDS-PAGE (4-12% gradient gels recommended).
- Transfer to PVDF membranes and probe with:
- M1-linkage specific monoclonal antibody (e.g., Millipore MABE1005)
- K63-linkage specific antibody (e.g., Millipore 05-1308)
- Pan-ubiquitin antibody (e.g., P4D1) as loading control
- Use secondary antibodies conjugated to fluorophores or HRP for detection.
Data Interpretation:
- M1-linear specific signal: Abolished by Otulin treatment but resistant to AMSH-LP
- K63-linked specific signal: Abolished by AMSH-LP treatment but resistant to Otulin
- Hybrid chains: Reduction in molecular weight after Otulin treatment with persistence of K63-signal, or vice versa
Diagram 3: Experimental workflow for distinguishing ubiquitin chain types using DUB specificity.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Essential Research Reagents for Studying Ubiquitin Chain Specificity
| Reagent Category | Specific Examples | Function/Application |
|---|---|---|
| Linkage-Specific Antibodies | Anti-linear ubiquitin (M1-specific); Anti-K63-linkage specific | Immunoblotting, immunofluorescence, immunohistochemistry for specific chain detection |
| Recombinant E3 Ligases | LUBAC complex (HOIP/HOIL-1/Sharpin); TRAF6; UBR5 | In vitro ubiquitination assays; reconstitution experiments in knockout cells |
| Deubiquitinases (DUBs) | Otulin (M1-specific); AMSH-LP (K63-specific) | Linkage specificity controls; validation of chain identity; cleavage of specific linkages |
| Ubiquitin-Binding Probes | Halo-NEMO; Tandem Ubiquitin Binding Entities (TUBEs) | Affinity capture of ubiquitinated proteins; pull-down assays |
| Cell Lines | LUBAC-deficient cells; TRAF6-deficient MEFs; NEMO-deficient cells | Genetic validation of specific pathway requirements; reconstitution studies |
| Activity-Based Probes | Ubiquitin vinyl sulfone; HA-Ub-VS | DUB activity profiling; identification of ubiquitin-interacting proteins |
| Mass Spectrometry Standards | DiGly antibody (K-ε-GG); SILAC ubiquitin | Proteomic identification of ubiquitination sites; linkage type determination |
The precise distinction between M1-linked, K63-linked, and branched ubiquitin chains is fundamental to understanding the complexity of immune signaling regulation. Each chain type possesses unique structural characteristics, is formed by specific enzymatic machinery, and fulfills distinct functions in cellular signaling pathways. The hierarchical relationship between these modifications—particularly the dependency of M1-linear chain formation on pre-existing K63-linked chains—reveals a sophisticated regulatory architecture that ensures precise control over immune and inflammatory responses. Continued advancement in detection methodologies and research reagents will further illuminate the nuanced functions of these ubiquitin chain types and their potential as therapeutic targets in immune-related diseases and cancer.
Navigating LUBAC Complex Stability and Auto-inhibition in Reconstitution Experiments
The Linear Ubiquitin Chain Assembly Complex (LUBAC) is a crucial regulator of inflammatory signaling, cell death, and immunity. For researchers aiming to dissect its functions, particularly in the context of M1-linked linear ubiquitin chain formation in immune signaling, successful reconstitution of the active complex is a fundamental prerequisite. This technical guide delves into the core challenges of LUBAC reconstitution, focusing on the structural basis of complex stability and the allosteric mechanisms of HOIP auto-inhibition. We provide detailed, experimentally-validated methodologies to overcome these hurdles, enabling the production of functionally active LUBAC for biochemical and structural studies.
LUBAC is the only known E3 ubiquitin ligase that catalyzes the formation of M1-linked linear ubiquitin chains, a critical post-translational modification in innate and adaptive immune signaling [4] [1]. The complex is a central signaling hub, regulating pathways downstream of receptors like the T cell receptor (TCR) and Toll-like receptors (TLR3) by modulating the NF-κB pathway and cell death decisions [72] [73]. The active complex is tripartite, consisting of:
- HOIP (RNF31): The catalytic subunit possessing linear ubiquitination activity.
- HOIL-1L (RBCK1): An accessory RBR E3 ligase that stabilizes the complex.
- SHARPIN: An adaptor protein crucial for complex integrity.
A primary obstacle in in vitro studies has been the inherent instability of individual subunits and the auto-inhibited state of full-length HOIP when expressed in isolation. This guide synthesizes current strategies to navigate these challenges, providing a roadmap for effective LUBAC reconstitution.
Structural Composition and Stability Mechanisms
The stability of the LUBAC holoenzyme is governed by specific, high-affinity interactions between its three core subunits. Understanding these interactions is the first step in designing a successful reconstitution strategy.
Table 1: Core Subunits of LUBAC and Their Domains
| Subunit | Key Domains | Primary Function in Complex Stability |
|---|---|---|
| HOIP | UBA, RBR, LDD, PUB | Catalytic core; provides scaffolding for HOIL-1L and SHARPIN binding via its UBA domain. |
| HOIL-1L | UBL, LTM, RBR | Forms a heterodimer with SHARPIN via LTM; its UBL domain binds HOIP's UBA domain. |
| SHARPIN | UBL, LTM, NZF | Forms a heterodimer with HOIL-1L via LTM; its UBL domain binds HOIP's UBA domain. |
The pivotal discovery is that the LUBAC-tethering motifs (LTMs) of HOIL-1L and SHARPIN fold into a single globular domain upon heterodimerization. This LTM-mediated dimerization is critical for the stability of the trimeric LUBAC complex [1]. Furthermore, the Ubiquitin-Associated (UBA) domain of HOIP serves as a binding platform for the Ubiquitin-Like (UBL) domains of both HOIL-1L and SHARPIN [38]. It is this network of UBA-UBL interactions, built upon the stable LTM heterodimer, that prevents HOIP destabilization and is essential for forming the active complex of 1:1:1 stoichiometry [3] [1].
Visualizing LUBAC Stability and Activation
The following diagram illustrates the domain architecture of LUBAC and the critical interactions that govern its stability and release from auto-inhibition.
Diagram 1: Domain architecture of LUBAC and critical stability interactions.
Overcoming HOIP Auto-inhibition
A major breakthrough in understanding LUBAC regulation was the finding that full-length HOIP is auto-inhibited by its own N-terminal domain. In vitro, the isolated HOIP RBR-LDD fragment displays constitutive linear ubiquitination activity, whereas the full-length protein is largely inactive [38]. This auto-inhibition is relieved by the binding of the UBL domains of HOIL-1L and SHARPIN to the UBA domain of HOIP [38]. This interaction induces conformational changes that allosterically rearrange the orientation between the UBA and RBR-LDD domains, facilitating E2 loading and promoting the catalytic activity of HOIP [38]. This mechanism underscores the biological rationale for the complex's tripartite nature: HOIL-1L and SHARPIN are not merely stabilizers but are essential allosteric activators.
Experimental Protocols for Reconstitution
Based on published successful structural and biochemical studies, the following protocol has been established for the reconstitution of active, stable LUBAC.
Co-expression and Tandem Affinity Purification
The most effective method to obtain a homogeneous, fully-assembled LUBAC complex is via co-expression in insect cells, which avoids the instability and improper folding associated with bacterial expression and assembly from individually purified subunits [3].
Table 2: Tandem Affinity Purification Protocol for LUBAC
| Step | Description | Critical Parameters & Tips |
|---|---|---|
| 1. Construct Design | Clone HOIP, His-tagged HOIL-1L, and Strep(II)-tagged SHARPIN into a baculovirus transfer vector. | Use a single vector for co-expression to ensure equimolar production of all subunits. |
| 2. Virus Generation | Generate recombinant baculovirus using the Bac-to-Bac system. | Amplify virus to a high titer (P2 or P3) for efficient infection. |
| 3. Cell Culture & Infection | Infect Trichoplusia ni (Hi5) or Spodoptera frugiperda (Sf9) cells at a density of 2-4 x 10^6 cells/mL with a high MOI. | Monitor cell viability; harvest cells 48-72 hours post-infection. |
| 4. Tandem Purification | Lyse cells and perform sequential affinity chromatography: first on a Strep-Tactin column (for SHARPIN), then on a Ni-NTA column (for HOIL-1L). | Use gentle lysis conditions. Include 1-2 mM DTT in buffers to preserve complex integrity. |
| 5. Size-Exclusion Chromatography (SEC) | Pool pure fractions and inject onto a Superose 6 Increase column pre-equilibrated with SEC buffer. | This step separates fully assembled LUBAC (eluting at ~0.94-0.97 mL on a Superose 6 3.2/300) from sub-complexes and aggregates. |
This co-expression strategy, followed by tandem affinity purification and SEC, yields a monodisperse complex, as verified by SDS-PAGE, immunoblotting, and negative-stain electron microscopy, which shows an elongated dumbbell structure [3].
Functional Validation of Reconstituted LUBAC
After purification, the linear ubiquitination activity of the reconstituted complex must be confirmed.
- In Vitro Ubiquitination Assay: Incubate ~100 nM of purified LUBAC with 100-200 nM E1 (UBA1), 1-2 µM E2 (UBE2L3), 10-20 µM ubiquitin, and an energy-regenerating system (1 mM ATP, 10 mM creatine phosphate, 10 U/mL creatine kinase) in reaction buffer (e.g., 50 mM Tris-HCl pH 7.5, 100 mM NaCl, 10 mM MgCl2) at 30°C.
- Analysis: Stop reactions at various time points (e.g., 0, 30, 60, 90 min) by adding SDS-PAGE sample buffer. Analyze by SDS-PAGE and immunoblotting using anti-linear ubiquitin antibodies (e.g., from MilliporeSigma or Life Technologies) to detect the formation of high-molecular-weight M1-linked ubiquitin chains.
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents for LUBAC Reconstitution and Analysis
| Reagent / Tool | Function / Application | Key Characteristics & Examples |
|---|---|---|
| Anti-Linear Ubiquitin Antibodies | Detection and quantification of M1-linked ubiquitin chains in immunoblotting. | Linkage-specific (e.g., MilliporeSigma 1:1000 dilution). Critical for validating LUBAC activity. |
| UBE2L3 (UbcH7) E2 Enzyme | The primary E2 conjugating enzyme that works with LUBAC. | Essential component of the in vitro ubiquitination assay. Commercially available (e.g., R&D Systems, Boston Biochem). |
| OTULIN DUB | Negative control; specifically disassembles linear ubiquitin chains. | Validates that observed chains are M1-linked. Commercially available (e.g., Life Technologies). |
| HOIP RBR-LDD Fragment | Positive control for catalytic activity, bypassing auto-inhibition. | Used to compare activity levels with the full trimeric complex. |
| Cysteine Mutant HOIP (C885A) | Catalytically dead negative control for LUBAC. | Mutations in the active site of HOIP's RING2 domain abolish activity. |
The faithful reconstitution of the LUBAC complex is a technically demanding but achievable goal. The key to success lies in respecting the complex's inherent structural biology: the critical dependence on co-expression to ensure proper folding and assembly, and the understanding that HOIL-1L and SHARPIN are essential co-activators, not just structural components. By employing the tandem affinity purification strategy in insect cells and rigorously validating output with linkage-specific tools, researchers can generate high-quality LUBAC for mechanistic studies. This, in turn, will accelerate our understanding of M1-linear ubiquitination in immune signaling and pave the way for developing therapeutics targeting this pathway in inflammation and cancer.
Optimizing Cell Lysis Conditions and Buffer Composition to Preserve Labile M1-Ubiquitin Modifications
The post-translational modification of proteins with M1-linked, or linear, ubiquitin chains is a crucial regulatory mechanism in innate immune signaling and inflammation [38]. Unlike canonical ubiquitin chains linked via lysine residues (e.g., K48, K63), linear ubiquitin is characterized by a head-to-tail linkage through the N-terminal methionine (M1) of the acceptor ubiquitin, forming a unique peptide bond [4] [74]. This specific topology is generated exclusively by the Linear Ubiquitin Chain Assembly Complex (LUBAC), a multi-protein complex comprising HOIP, HOIL-1L, and SHARPIN [4] [38]. LUBAC-mediated linear ubiquitination plays a non-redundant role in fine-tuning the output of key signaling pathways, including those triggered by Tumor Necrosis Factor (TNF), interleukin-1β (IL-1β), and Toll-like receptors (TLRs) [75] [74]. The primary function of M1 chains in these pathways is not to trigger proteasomal degradation but to act as scaffolds for the assembly and activation of signaling complexes, most notably those leading to the activation of the transcription factor NF-κB [4] [75].
Given their central role in orchestrating the immune response, the accurate detection and quantification of M1-ubiquitin chains are paramount. However, these modifications are notably labile and transient, making their preservation during cell lysis a significant technical challenge [38]. The dynamic nature of ubiquitination is maintained by the opposing actions of writers (E3 ligases like LUBAC) and erasers—deubiquitinases (DUBs) such as OTULIN and CYLD, which specifically target and disassemble M1-linked chains [4]. If not properly controlled during sample preparation, the activities of these DUBs, as well as proteasomal degradation, can rapidly erase the ubiquitin signature a researcher aims to capture [76]. Therefore, optimizing lysis conditions is not merely a preparatory step but a critical determinant for obtaining biologically relevant data on linear ubiquitination in the context of immune signaling and drug discovery.
Key Challenges in Preserving M1-Ubiquitin Modifications
The successful capture of endogenous M1-ubiquitin chains is fraught with difficulties stemming from the cellular machinery itself and the intrinsic properties of the ubiquitin code.
- DUB Activity: The deubiquitinases OTULIN and CYLD are highly specific for cleaving M1-linked ubiquitin chains [4]. These enzymes remain active in standard cell lysates and can rapidly dismantle linear ubiquitin chains on substrates like RIPK1 and NEMO within minutes of cell disruption, leading to a profound loss of signal [4] [76].
- Proteasomal Degradation: While M1 chains themselves are generally non-proteolytic, other ubiquitin linkages (e.g., K48, K11) that may coexist on the same protein or in the same complex can target it for proteasomal degradation [4] [76]. Inhibiting the proteasome is therefore essential to prevent the indirect loss of proteins modified with mixed or branched ubiquitin chains.
- Linkage Lability: Research indicates that K63 and M1 linkages are particularly sensitive to degradation during sample preparation. Standard concentrations of the DUB inhibitor N-ethylmaleimide (NEM) (5-10 mM) are insufficient; preserving K63 linkages requires up to 10 times higher concentrations of NEM, suggesting similar stringent conditions may be needed for the equally labile M1 chains [76].
- Antibody Recognition Issues: Many commercial anti-ubiquitin antibodies do not recognize all linkage types equally. For instance, some widely used antibodies poorly detect M1-linkages compared to K48 or K63 chains, which can lead to a significant underestimation of M1-ubiquitin levels in western blot analyses [76].
Optimized Lysis Buffer Composition for M1-Ubiquitin Preservation
The foundation for preserving M1-ubiquitin modifications is a tailored lysis buffer designed to simultaneously inactivate decomplexing enzymes and maintain the native state of ubiquitinated proteins. The following table provides a detailed breakdown of the essential components and their optimized concentrations.
Table 1: Key Components of an Optimized Lysis Buffer for M1-Ubiquitin Preservation
| Component | Recommended Concentration | Functional Role | Critical Considerations |
|---|---|---|---|
| Buffering Agent (e.g., Tris-HCl) | 20-50 mM, pH 7.4-7.6 [77] | Maintains physiological pH for protein stability [78]. | pH is critical; avoid acidic conditions that can destabilize ubiquitin complexes. |
| Detergent (e.g., NP-40, Triton X-100) | 1% [79] [77] | Solubilizes membranes and extracts proteins under non-denaturing conditions. | Avoid SDS in initial lysis if co-immunoprecipitation is planned, as it denatures proteins [79]. |
| Salts (e.g., NaCl) | 150 mM [77] | Maintains ionic strength and prevents non-specific protein aggregation. | |
| DUB Inhibitor (N-Ethylmaleimide, NEM) | 50-100 mM [76] | Irreversibly alkylates catalytic cysteine residues of DUBs like OTULIN and CYLD [80]. | Crucial for M1 preservation. Prepare fresh in ethanol before use. High concentration is required [80] [76]. |
| Proteasome Inhibitor (e.g., MG132) | 10-20 µM | Inhibits the 26S proteasome, preventing degradation of proteins marked with degradative ubiquitin chains. | Avoid prolonged pre-lysis treatment (>12h) to prevent stress-induced ubiquitination [76]. |
| Chelating Agents (EDTA, EGTA) | 1-5 mM [77] | Chelates metal ions, inhibiting some metal-dependent proteases and DUBs. | |
| Additional Protease Inhibitors | Cocktail (e.g., Leupeptin 1 µg/mL [77]) | Broad-spectrum inhibition of serine, cysteine, and metalloproteases. | Use commercially available cocktails for consistency. |
Recommended Buffer Formulations
Based on the components above, here are two proven starting formulations for lysis buffer:
Formulation A: General Purpose M1-Preservation Lysis Buffer This buffer is ideal for most immunoprecipitation and western blot applications.
- 20 mM Tris-HCl, pH 7.5 [77]
- 150 mM NaCl [77]
- 1% Triton X-100 [77]
- 1 mM Na₂EDTA & 1 mM EGTA [77]
- 50 mM NEM (freshly added) [76]
- 10 µM MG132
- 1 µg/mL Leupeptin [77] or commercial protease inhibitor cocktail
Formulation B: DiGLY Proteomics Lysis Buffer For mass spectrometry-based ubiquitomics, a denaturing buffer is preferred to instantly inactivate all enzymes.
- 8 M Urea [80]
- 150 mM NaCl [80]
- 50 mM Tris-HCl, pH 8.0 [80]
- 5 mM NEM (freshly added) [80] Note: Concentration can be optimized upwards.
- 1 mM NaF, 1 mM β-glycerophosphate, 1 mM Na₃VO₄ (phosphatase inhibitors) [80]
- Protease inhibitor cocktail [80]
Detailed Experimental Workflow for Cell Lysis and Sample Preparation
A meticulous, cold-based workflow is essential from the moment of cell harvesting to ensure the integrity of M1-ubiquitin modifications.
Pre-Lysis Considerations
- Inhibitor Preparation: Pre-chill all buffers and centrifuges to 4°C. Prepare stock solutions of NEM and MG132 and add them to the ice-cold lysis buffer immediately before use [80] [76] [77].
- Cell Handling: For adherent cells, wash the monolayer once with ice-cold Phosphate-Buffered Saline (PBS) to remove residual media. For suspension cells, pellet and wash with PBS [77].
Lysis Procedure
- Lysate Generation: Add the appropriate volume of chilled lysis buffer directly to the cell pellet or plate (e.g., 400 µL per 10⁷ cells or a 10 cm dish) [77].
- Incubation: Incubate on ice for 5-15 minutes with gentle agitation [77].
- Mechanical Disruption: For adherent cells, scrape them from the plate while immersed in buffer. For tissues, mechanical homogenization (e.g., Dounce homogenizer) is necessary [79] [77].
- Clarification: Transfer the lysate to a microcentrifuge tube. Sonicate briefly (e.g., 3-5 pulses of 5-10 seconds on ice) to shear DNA and reduce viscosity [77].
- Clearing: Centrifuge at 14,000 x g for 10-15 minutes at 4°C to pellet insoluble debris [77].
- Storage: Immediately transfer the clarified supernatant (whole cell extract) to a new tube. Aliquot and flash-freeze in liquid nitrogen for long-term storage at -80°C. Avoid multiple freeze-thaw cycles.
Downstream Detection and Analysis of M1-Ubiquitin Modifications
Western Blotting Optimization
The high molecular weight of polyubiquitinated proteins requires specific electrophoretic conditions for clear resolution.
- Gel Selection: For resolving large ubiquitin chains (up to 20+ units), use 8% Tris-glycine SDS-PAGE gels. For better separation of smaller chains (e.g., di-ubiquitin), 12% gels are preferable [76].
- Buffer Systems: Use MOPS-based running buffer for optimal resolution of long chains (>8 ubiquitin units) and MES-based buffer for smaller chains (2-5 units) [76].
- Membrane and Transfer: PVDF membranes (0.2 µm pore size) provide superior signal strength for ubiquitin detection. Perform transfers at a constant 30 V for 2.5 hours to prevent incomplete transfer or unfolding of ubiquitin chains, which can impair antibody recognition [76].
- Antibody Validation: Carefully select and validate antibodies. Many common anti-ubiquitin antibodies (e.g., from Dako, Cell Signaling Technology) have varying affinities for different linkages and may poorly recognize M1 chains. Where possible, use linkage-specific antibodies or ubiquitin-binding domains (UBDs) for pull-down [76].
DiGLY Proteomics for Site-Specific Mapping
For system-wide identification of ubiquitination sites, including M1-linked substrates, diGLY proteomics is the gold standard. This method involves:
- Cell Lysis: Using a denaturing buffer like Formulation B to instantly freeze ubiquitination states [80].
- Trypsin Digestion: Trypsin cleaves proteins after lysine and arginine, but leaves a signature di-glycine (diGLY) remnant on the modified lysine of the substrate [80].
- Immunoaffinity Enrichment: diGLY-modified peptides are enriched from the complex peptide mixture using specific anti-diGLY antibodies [80].
- Mass Spectrometry: The enriched peptides are identified and quantified by LC-MS/MS, allowing for the mapping of ubiquitination sites and the relative abundance of ubiquitinated proteins across conditions [80].
Table 2: Essential Research Reagents for M1-Ubiquitin Studies
| Reagent Category | Specific Examples | Function in Research |
|---|---|---|
| LUBAC Components | HOIP (RNF31), HOIL-1L (RBCK1), SHARPIN [4] [38] | Targets for genetic manipulation (KO, KD) to study LUBAC-specific functions. |
| M1-Specific DUBs | OTULIN, CYLD [4] | Critical controls and targets for validating the specificity of M1-chain detection. |
| Inhibitors | N-Ethylmaleimide (NEM), MG132 [80] [76] | Essential additives in lysis buffer for preserving ubiquitin chains. |
| Linkage-Specific Tools | Anti-linear ubiquitin antibody (e.g., Ubiquigent), TUBE assays [76] [38] | Key reagents for the specific detection and pull-down of M1-ubiquitin chains. |
| diGLY Proteomics Kits | PTMScan Ubiquitin Remnant Motif (K-Ɛ-GG) Kit [80] | For systematic, site-specific identification of ubiquitination sites by mass spectrometry. |
The Signaling Context: M1-Ubiquitin in TNF Receptor Signaling
The TNF receptor 1 (TNFR1) pathway provides a canonical example of where optimized lysis is critical for studying M1-ubiquitin function. Upon TNF binding, TNFR1 forms a membrane-associated signaling complex (Complex I) containing TRADD, TRAF2, RIPK1, and cIAP1/2 [74]. cIAPs mediate K63-linked ubiquitination of RIPK1, which serves as a platform for the recruitment of LUBAC. LUBAC then generates M1-linked ubiquitin chains on components of the Complex I, including RIPK1 and NEMO (IKKγ) [4] [75] [74]. These M1 chains act as a specific beacon for the recruitment of the IKK complex through the high-affinity binding of NEMO to linear ubiquitin. This recruitment is essential for the full activation of IKK and the subsequent NF-κB signaling cascade, which promotes cell survival and inflammatory gene expression [75] [74]. Without optimized lysis conditions that preserve these labile M1 and K63 chains, the composition and molecular architecture of this critical signaling complex cannot be accurately studied.
M1-linked linear ubiquitination is a distinct post-translational modification where ubiquitin molecules form head-to-tail chains through a peptide bond between the C-terminal glycine of one ubiquitin and the N-terminal methionine (M1) of the next [4] [74]. Unlike canonical K48-linked chains that target proteins for proteasomal degradation, M1-linked chains primarily function as scaffolds for signal transduction, playing pivotal roles in regulating immune signaling, cell death, and inflammation [81] [82]. The Linear Ubiquitin Chain Assembly Complex (LUBAC), comprised of HOIP, HOIL-1L, and SHARPIN, serves as the sole E3 ligase capable of catalyzing linear ubiquitin chain formation [4] [82] [2]. This modification is dynamically regulated by deubiquitinases (DUBs) OTULIN and CYLD, which specifically disassemble M1-linked chains [4] [82]. The critical importance of this pathway is demonstrated by the severe phenotypes observed in various knockout models, where disruption of LUBAC components or regulators leads to profound immune dysregulation, altered cell death responses, and developmental defects [4] [82] [2].
The Core Machinery of M1-Linear Ubiquitination
Writers, Erasers, and Readers of the Linear Ubiquitin Code
The linear ubiquitination system operates through a coordinated interplay of enzymatic writers, erasers, and specialized readers that interpret the ubiquitin code [82] [2]. The precise regulation of this system is essential for maintaining cellular homeostasis, particularly in immune cells where signaling must be robust yet tightly controlled to prevent pathological inflammation [32] [81].
Table 1: Core Components of the M1-Linear Ubiquitination System
| Component Type | Molecular Elements | Primary Functions | Key Domains/Motifs |
|---|---|---|---|
| Writers | LUBAC complex (HOIP, HOIL-1L, SHARPIN) | Catalyzes M1-linked ubiquitin chain formation | HOIP: RBR, LDD, UBA, PUB; HOIL-1L: RBR, UBL; SHARPIN: UBL, LTM |
| Erasers | OTULIN | Specifically hydrolyzes linear ubiquitin chains | OTU domain, PIM motif |
| CYLD | Removes both K63-linked and linear ubiquitin chains | USP domain, CAP-Gly domains | |
| Readers | NEMO (IKKγ) | Binds linear chains to activate IKK complex and NF-κB signaling | UBAN/CoZi domain |
| A20 (TNFAIP3) | Recognizes linear chains to suppress NF-κB signaling | OTU domain, ZnF7 | |
| ABIN proteins | Bind linear chains with functions in signal regulation | UBAN domain |
Structural and Functional Basis of LUBAC
HOIP serves as the catalytic core of LUBAC, containing a unique linear ubiquitin chain determining domain (LDD) that positions acceptor ubiquitins for M1-linkage formation [82] [2] [38]. The RING-between-RING (RBR) domain of HOIP employs a hybrid mechanism, first accepting ubiquitin from an E2 enzyme (typically UBE2L3) via a transient thioester intermediate before transferring it to the substrate [38]. HOIP's catalytic activity is autoinhibited in isolation but is released upon binding of HOIL-1L and SHARPIN through their ubiquitin-like (UBL) domains interacting with HOIP's ubiquitin-associated (UBA) domain [2]. This structural arrangement ensures that LUBAC functions exclusively as a heterotrimeric complex, providing an additional layer of regulatory control. HOIL-1L itself possesses E3 ligase activity and can monoubiquitinate LUBAC subunits, potentially attenuating complex function, while SHARPIN primarily serves as a structural stabilizer [82] [2].
Phenotypic Analysis of M1-Ubiquitination Knockout Models
Genetic ablation of components in the linear ubiquitination pathway produces distinctive phenotypic outcomes across different model systems, revealing both shared and unique functions of each element. The quantitative data from these models provides crucial insights into the essential nature of proper M1-ubiquitination regulation for cellular and organismal homeostasis.
Table 2: Comprehensive Phenotypic Outcomes in M1-Ubiquitination Pathway Knockout Models
| Target Gene | Model System | Key Cellular Phenotypes | Organismal/Pathological Outcomes | Signaling Pathways Affected |
|---|---|---|---|---|
| HOIP | Mouse knockout | Embryonic lethality by E10.5-12.5; endothelial cell death | Impaired vascular development; defective lymphorganogenesis | TNFα-induced NF-κB activation severely impaired; TNFR1-mediated apoptosis enhanced |
| HOIL-1L | Mouse knockout | Impaired NF-κB activation in fibroblasts and B cells; no embryonic lethality | Reduced B cell numbers and antibody responses; protection from TNF-induced shock | Partial defect in TNFα and IL-1β signaling; CD40 and TLR signaling impaired |
| SHARPIN | cpdm mice (natural mutant) | Chronic proliferation dermatitis; keratinocyte hyperproliferation and death | Severe skin inflammation; lymphoid organ defects; immune cell infiltration | TNFα-induced NF-κB activation impaired; TNFR1-mediated apoptosis enhanced; spontaneous inflammation |
| OTULIN | Mouse knockout (Gumby) | Embryonic lethality by E14.5; angiogenic defects | Impaired vasculogenesis; Wnt signaling defects | TNF-induced NF-κB hyperactivation (due to LUBAC auto-ubiquitination prevention) |
| OTULIN | Human mutations (ORAS) | Systemic autoinflammation in patients | Fever, rash, joint inflammation, diarrhea, lipodystrophy | Elevated inflammatory cytokines; leukocytosis |
| CYLD | Mouse knockout | No major developmental defects | Enhanced susceptibility to colitis-associated cancer; enhanced inflammation upon challenge | Enhanced NF-κB and JNK activation in response to TLR and TNF stimulation |
Interpretation of Knockout Phenotypes and Compensatory Mechanisms
The embryonic lethality observed in HOIP-deficient mice underscores the fundamental role of linear ubiquitination in developmental processes, particularly in vascular and immune system formation [4] [82]. This severe phenotype contrasts with the viability of HOIL-1L knockout mice, suggesting that HOIP retains partial functionality even in the absence of its binding partner, though with significantly compromised activity across multiple signaling pathways [82]. The cpdm (chronic proliferative dermatitis) phenotype in SHARPIN-deficient mice reveals the critical importance of LUBAC integrity for preventing spontaneous inflammation and maintaining epidermal homeostasis [4] [74]. Interestingly, the different phenotypic outcomes between OTULIN and CYLD knockout models highlight the distinct biological functions of these deubiquitinases despite their shared ability to cleave linear ubiquitin chains. OTULIN deficiency leads to embryonic lethality or severe autoinflammation, while CYLD knockout mice develop relatively normally but exhibit enhanced inflammatory responses to challenges, indicating that OTULIN plays a non-redundant role in preventing aberrant LUBAC activation and signaling dysregulation [82] [2].
Experimental Approaches for Analyzing M1-Ubiquitination Defects
Methodologies for Assessing Linear Ubiquitination in Cellular Models
Investigating M1-ubiquitination requires specialized methodologies capable of specifically detecting this unique ubiquitin linkage type amid the complex background of other ubiquitin modifications. The following experimental protocols represent key approaches for comprehensive analysis of linear ubiquitination defects in knockout models:
1. Immunoblotting with Linear Ubiquitin-Specific Antibodies
- Procedure: Resolve protein extracts from wild-type and knockout cells by SDS-PAGE. Transfer to membranes and probe with monoclonal antibodies specifically recognizing M1-linked ubiquitin chains (e.g., Anti-linear ubiquitin [1E3]).
- Key Applications: Detect endogenous linear ubiquitin conjugates; monitor changes in global M1-ubiquitination levels in knockout models; verify LUBAC activity in reconstitution experiments.
- Technical Considerations: Include samples treated with OTULIN (linear chain-specific DUB) as a specificity control. Combine with immunoprecipitation of specific proteins to assess their linear ubiquitination status.
2. Immunofluorescence and Proximity Ligation Assay (PLA)
- Procedure: Fix cells, permeabilize, and incubate with primary antibodies against target proteins and linear ubiquitin. For PLA, use species-specific secondary antibodies with connector oligonucleotides to visualize proximal interactions.
- Key Applications: Visualize subcellular localization of linear ubiquitin chains; detect protein complexes containing M1-ubiquitinated targets; validate recruitment of LUBAC to specific signaling complexes.
- Technical Considerations: Optimize fixation conditions to preserve ubiquitin chain architecture. Include appropriate controls (DUB pretreatment, competition with recombinant ubiquitin chains) to verify specificity.
3. Tandem Ubiquitin Binding Entity (TUBE) Pulldown with Linkage-Specific Elution
- Procedure: Incubate cell lysates with Tandem Ubiquitin Binding Entities (TUBEs) to capture polyubiquitinated proteins. Wash extensively, then elute with buffers containing specific diubiquitin linkages (K48-, K63-, or M1-linked) to competitively release subsets of ubiquitinated proteins.
- Key Applications: Isolate and quantify proteins modified with specific ubiquitin chain types; identify novel linear ubiquitination substrates; monitor chain-type dynamics in signaling pathways.
- Technical Considerations: Use recombinantly expressed and purified diubiquitins for competitive elution. Validate specificity with linkage-specific DUBs (OTULIN for M1-linkages).
Functional Assays for Pathway Analysis in Knockout Models
1. NF-κB Luciferase Reporter Assay
- Procedure: Transfect wild-type and knockout cells with NF-κB-responsive luciferase reporter construct and stimulation controls. Treat cells with pathway-specific agonists (TNFα, IL-1β, LPS) for varying durations. Measure luciferase activity as a readout of NF-κB activation.
- Key Applications: Quantitatively assess the impact of M1-ubiquitination defects on NF-κB signaling amplitude and kinetics; test functional rescue with LUBAC component reconstitution.
- Technical Considerations: Include minimal promoter controls to normalize for transfection efficiency and non-specific effects. Use pharmacological inhibitors (IKK inhibitors, proteasome inhibitors) to validate pathway specificity.
2. Cytokine Profiling by ELISA and Multiplex Assays
- Procedure: Stimulate wild-type and knockout macrophages, fibroblasts, or other relevant cell types with innate immune agonists (LPS, TNFα, cyclic dinucleotides). Collect supernatants at various time points and quantify cytokine/chemokine production using ELISA or bead-based multiplex arrays.
- Key Applications: Characterize the impact of linear ubiquitination defects on inflammatory output; identify specific cytokine classes most dependent on M1-ubiquitination (e.g., type I IFNs vs. proinflammatory cytokines).
- Technical Considerations: Include both early (IL-6, TNFα) and late (IFN-β, IL-12) cytokines to distinguish primary and secondary signaling defects. Perform dose-response curves to reveal potential signaling thresholds.
3. Co-immunoprecipitation and Protein Complex Analysis
- Procedure: Generate lysates from stimulated wild-type and knockout cells under non-denaturing conditions. Immunoprecipitate signaling complex components (TNFR1, NEMO, RIPK1). Analyze co-precipitating proteins by immunoblotting for LUBAC components, ubiquitin chains, and downstream effectors.
- Key Applications: Determine composition and dynamics of signalosomes in the presence and absence of linear ubiquitination; identify specific protein interactions dependent on M1-chains.
- Technical Considerations: Use crosslinking for transient interactions. Combine with mass spectrometry for unbiased identification of complex components.
Signaling Pathway Visualization and Molecular Relationships
The intricate relationships between M1-ubiquitination components and their positions within immune signaling pathways can be visualized through the following diagrams:
M1-Ubiquitination in TNF Signaling and Knockout Phenotypes
LUBAC Complex Assembly and Regulatory Mechanisms
Essential Research Reagents and Experimental Tools
The study of M1-ubiquitination requires specialized reagents that specifically target components of this pathway. The following toolkit represents essential resources for investigating linear ubiquitination in knockout models and experimental systems.
Table 3: Research Reagent Solutions for M1-Ubiquitination Studies
| Reagent Category | Specific Examples | Research Applications | Technical Considerations |
|---|---|---|---|
| Linkage-Specific Antibodies | Anti-linear ubiquitin (1E3, Millipore); Anti-K63 ubiquitin; Anti-K48 ubiquitin | Immunoblotting, immunofluorescence, immunohistochemistry; Distinguish M1-linkages from other ubiquitin modifications | Validate specificity with linkage-specific DUBs (OTULIN for M1); Optimize conditions to preserve chain architecture |
| Recombinant LUBAC Components | Recombinant trimeric LUBAC complex; Individual subunits (HOIP, HOIL-1L, SHARPIN) | In vitro ubiquitination assays; Reconstitution studies in knockout cells; Structural studies | Co-express subunits for proper complex formation; Include catalytically inactive mutants (HOIP C885A) as controls |
| Activity-Based Probes | Linear ubiquitin-specific DUB probes; TUBE (Tandem Ubiquitin Binding Entities) | Monitor DUB activity in cell extracts; Enrich and identify M1-ubiquitinated proteins | Use linkage-specific diubiquitin standards for validation; Combine with mass spectrometry for substrate identification |
| Cell Line Models | HOIP-/- MEFs; SHARPIN-deficient cpdm cells; OTULIN-/- macrophages | Pathway analysis in controlled genetic backgrounds; Signaling studies without compensation | Verify knockout status with multiple methods; Monitor potential adaptive changes in parallel pathways |
| Animal Models | HOIP conditional knockout mice; SHARPIN cpdm mice; OTULIN Gumby mice | In vivo studies of immune function, development, inflammation | Consider cell-type specific functions with conditional models; Account for potential embryonic lethality with inducible systems |
Concluding Perspectives and Future Directions
The comprehensive analysis of M1-ubiquitination knockout models reveals the critical importance of this modification in maintaining immunological balance and cellular homeostasis. The distinct yet overlapping phenotypes observed in HOIP, HOIL-1L, SHARPIN, OTULIN, and CYLD deficient systems demonstrate both the core function of linear ubiquitination in NF-κB activation and its pathway-specific roles in regulating cell death, inflammation, and development. The experimental frameworks outlined here provide robust methodologies for interrogating linear ubiquitination defects across cellular and animal models, enabling researchers to precisely dissect the molecular consequences of disrupting this key signaling pathway. As research in this field advances, the growing understanding of M1-ubiquitination will continue to illuminate fundamental mechanisms of immune regulation and identify novel therapeutic opportunities for inflammatory diseases, immune disorders, and cancer.
Context and Specificity: Validating M1-Ubiquitin Functions Against the Broader Ubiquitin Landscape
Ubiquitination is a crucial post-translational modification that regulates a vast array of cellular processes, with particular significance in innate immune signaling. The functional outcome of ubiquitination is largely determined by the topology of the polyubiquitin chains formed. Among the eight possible linkage types, M1-linked (linear), K63-linked, and K48-linked chains represent three distinct archetypes with specialized roles in immune regulation. K48-linked chains primarily target proteins for proteasomal degradation, while K63-linked and M1-linked chains function as scaffolds for signal activation and amplification. Understanding the functional divergence between these chain types is essential for deciphering the complexity of immune responses and developing targeted therapeutic interventions. This review examines the specialized roles, signaling mechanisms, and functional interplay between M1, K63, and K48 ubiquitin chains in innate immunity, with particular focus on recent advances in linear ubiquitination research.
Structural and Functional Characteristics of Ubiquitin Chains
Table 1: Core Functional Characteristics of Major Ubiquitin Chain Types
| Feature | K48-Linked Chains | K63-Linked Chains | M1-Linked (Linear) Chains |
|---|---|---|---|
| Primary Function | Proteasomal degradation [83] [75] | Non-degradative signaling & complex assembly [75] [43] | NF-κB activation & inflammation regulation [31] [4] |
| Chain Conformation | Compact structure [83] | Extended, open structure [83] | Linear, rigid structure [38] |
| Key E3 Ligases | HUWE1, TRAF6 (branched) [84] | TRAF6, cIAPs [75] [43] | LUBAC (HOIP/HOIL-1L/SHARPIN) [4] [38] |
| Specialized DUBs | Multiple | CYLD, A20 [75] [84] | OTULIN, CYLD [4] [38] |
| Role in Cell Death | Indirect via degradation | Limited regulation | Direct regulation of apoptosis, necroptosis, pyroptosis [4] |
The structural and functional divergence between ubiquitin chain types originates from their distinct biophysical properties and recognition by specialized effector proteins. K48-linked chains adopt a compact conformation that facilitates recognition by proteasomal receptors, effectively marking substrate proteins for degradation [83]. In contrast, K63-linked chains exhibit an extended, open conformation that enables them to function as scaffolds for assembling signaling complexes without triggering degradation [83]. M1-linked linear chains form a unique rigid, linear structure that creates specific binding platforms for components of inflammatory signaling pathways, particularly in NF-κB activation [38].
The synthesis of each chain type is catalyzed by specialized E3 ligase complexes. The Linear Ubiquitin Chain Assembly Complex (LUBAC), composed of HOIP, HOIL-1L, and SHARPIN, serves as the exclusive E3 ligase for M1-linked chain formation [4] [38]. HOIP contains the catalytically essential Linear Ubiquitin chain Determining Domain (LDD) that specifically positions acceptor ubiquitins for M1-linkage [38]. K63-linked chains are synthesized by various E3 ligases including TRAF6 and XIAP in immune signaling contexts [75] [43], while K48-linked chains are generated by multiple E3s including HUWE1, which can collaborate with TRAF6 to form branched K48-K63 chains [84].
Deubiquitinating enzymes (DUBs) provide specificity in chain disassembly, with OTULIN and CYLD showing preference for M1-linked chains [4] [38], while CYLD and A20 also target K63-linked chains in NF-κB signaling pathways [75] [84]. This sophisticated interplay between writers, readers, and erasers of the ubiquitin code enables precise temporal and spatial control of innate immune signaling.
M1-Linked Linear Ubiquitin Chains in Immune Signaling
The Linear Ubiquitin Assembly Complex (LUBAC)
LUBAC represents the sole E3 ubiquitin ligase capable of generating M1-linked linear ubiquitin chains [4]. This multi-subunit complex consists of three core components: HOIP (57kDa, RNF31), which contains the catalytic RBR-LDD domain; HOIL-1L (123kDa, RBCK1), an RBR E3 ligase that stabilizes the complex; and SHARPIN (40kDa), which facilitates complex assembly [4] [38]. The interaction between ubiquitin-associated (UBA) domains of HOIP and ubiquitin-like (UBL) domains of HOIL-1L and SHARPIN releases HOIP from autoinhibition, enabling catalytic activity [38]. LUBAC function is tightly regulated by the deubiquitinases OTULIN and CYLD, which remove M1-linked chains and prevent excessive inflammatory signaling [4] [38].
Signaling Pathways Regulated by Linear Ubiquitination
Table 2: Key Signaling Pathways Regulated by M1-Linked Ubiquitin Chains
| Signaling Pathway | LUBAC Substrate | Functional Outcome | Biological Significance |
|---|---|---|---|
| NF-κB Signaling | NEMO/IKK complex [75] | Enhances IKK recruitment & activation | Pro-inflammatory gene expression |
| TNFR Signaling | RIPK1 [75] | Precedes K63 ubiquitination | Cell survival vs. death decisions |
| STING Pathway | STING [31] | Promotes NF-κB & IRF3 signaling | Antiviral response & interferon production |
| NLRP3 Inflammasome | NLRP3 components [75] | Regulates inflammasome assembly | IL-1β maturation & pyroptosis |
| Cell Death Regulation | Multiple (RIPK1, etc.) [4] | Modulates apoptosis, necroptosis | Tissue homeostasis & inflammation |
Recent research has illuminated the crucial role of linear ubiquitination in STING-mediated innate immunity. Fischer et al. (2025) demonstrated that STING activation induces HOIP-mediated synthesis of M1 ubiquitin chains that stimulate both NF-κB and IRF3 signaling pathways in human monocytes and mouse macrophages [31]. This M1-chain formation occurs on LC3B-associated Golgi membranes where LUBAC recruitment facilitates NF-κB activation independently of the canonical STING-IRF3 pathway. Notably, loss of HOIP prevents M1-chain formation and significantly reduces STING-induced signaling without affecting STING activation itself, highlighting the essential role of linear ubiquitination in this immune pathway [31].
In TNF receptor signaling, LUBAC-mediated linear ubiquitination of components within the TNFR complex I, particularly RIPK1 and NEMO, creates a platform for IKK complex recruitment and activation [75]. This M1-ubiquitination precedes and facilitates subsequent K63-chain formation, ultimately leading to IκBα phosphorylation, K48-linked ubiquitination, and proteasomal degradation, which releases NF-κB for nuclear translocation and pro-inflammatory gene expression [75]. The strategic positioning of M1-chains early in signaling cascades enables them to function as master regulators of inflammatory response amplitude and duration.
K63-Linked Ubiquitin Chains in Innate Immune Signaling
K63-linked ubiquitin chains serve as critical scaffolds for the assembly and activation of signaling complexes in multiple innate immune pathways. In TLR/IL-1R signaling, K63 ubiquitination of TRAF6 and other adaptor proteins creates platforms that recruit TAK1 complexes through ubiquitin-binding domains [75]. This recruitment initiates a kinase cascade leading to IKK and NF-κB activation. Similarly, in NOD2 signaling, bacterial muramyl dipeptide triggers RIPK2 ubiquitination with K63-linked chains by E3 ligases including XIAP, facilitating the recruitment of TAK1 and IKK complexes [43].
The functional significance of K63 chains extends beyond mere scaffolding. Recent evidence indicates that K63 ubiquitination can undergo functional reprogramming through branching. During IL-1β signaling, HUWE1 cooperates with TRAF6 to generate K48 branches on K63 chains, forming K48-K63 branched ubiquitin chains that protect K63 linkages from CYLD-mediated deubiquitination while maintaining recognition by TAB2 [84]. This branching mechanism amplifies and sustains NF-κB signaling beyond what would be possible with homogeneous K63 chains alone, demonstrating how linkage mixing can create more sophisticated regulatory outcomes.
K48-Linked Ubiquitin Chains and Immune Regulation
While traditionally associated with protein degradation, K48-linked ubiquitin chains participate in innate immune signaling through both degradative and non-degradative mechanisms. The canonical function involves targeting inhibitory proteins for proteasomal degradation, as exemplified by IκBα degradation in NF-κB signaling, which releases the transcription factor for nuclear translocation [75]. This degradation creates a negative feedback loop that limits the duration of NF-κB activation.
However, emerging research reveals more nuanced roles for K48 linkages in immune regulation. When incorporated as branches on K63-linked chains, K48 linkages can modulate signal duration without causing complete substrate degradation. In the aforementioned branched K48-K63 chains, the K48 component protects the chain from deubiquitination by CYLD while remaining accessible to TAB2 recognition, demonstrating how K48 linkages can influence signaling outcomes beyond protein degradation [84]. Additionally, K48-linked ubiquitination of specific immune regulators like TRIM27 contributes to sepsis pathogenesis by targeting PPARγ for degradation and exacerbating oxidative stress in lung tissues [75].
Branched and Mixed Ubiquitin Chains
Table 3: Characteristics of Branched Ubiquitin Chains in Immune Signaling
| Branched Chain Type | Forming E3 Ligases | Signaling Pathway | Functional Consequence |
|---|---|---|---|
| K48-K63 Branched | HUWE1 + TRAF6 [84] | IL-1R/NF-κB signaling | Protects K63 chains from CYLD, amplifies signaling [84] |
| K48-K63 Branched | UBR5 + ITCH [85] | Apoptotic regulation | Converts non-degradative to degradative signal [85] |
| K11-K48 Branched | APC/C (UBE2C+UBE2S) [85] | Cell cycle regulation | Targets mitotic regulators for degradation |
| K29-K48 Branched | Ufd4 + Ufd2 [85] | Ubiquitin fusion degradation | Quality control pathway |
Beyond homogeneous chains, branched ubiquitin chains represent a sophisticated mechanism for integrating multiple regulatory signals within a single ubiquitin polymer. These branched architectures significantly expand the coding potential of ubiquitin signaling by combining the properties of different linkage types [85]. The synthesis of branched chains frequently involves collaboration between E3 ligases with distinct linkage specificities, such as the partnership between TRAF6 (K63-specific) and HUWE1 (K48-specific) in NF-κB signaling [84].
Branched chains can serve distinct functions based on their architecture and synthesis order. In some contexts, branching enables signal conversion from non-degradative to degradative, as seen when UBR5 adds K48 linkages to K63 chains initially synthesized by ITCH on TXNIP, resulting in proteasomal degradation of this apoptotic regulator [85]. Alternatively, branching can create protective structures that resist deubiquitination, as demonstrated by K48-K63 branched chains that shield K63 linkages from CYLD-mediated disassembly while maintaining recognition by signaling components like TAB2 [84].
The emerging understanding of branched ubiquitination reveals an additional layer of complexity in immune regulation, where the integration of multiple linkage types within single polymers enables more precise control over signaling amplitude, duration, and specificity than would be possible with homogeneous chains alone.
Experimental Approaches and Research Tools
Methodologies for Studying Ubiquitin Chain Function
Advanced methodologies have been developed to decipher the complex functions of different ubiquitin chain types in innate immune signaling. Tandem Ubiquitin Binding Entities (TUBEs) represent a powerful tool for linkage-specific enrichment and detection of ubiquitinated proteins [43]. These engineered affinity reagents with nanomolar affinities for specific polyubiquitin linkages enable researchers to capture endogenous ubiquitination events without requiring genetic manipulation of the ubiquitin system.
A recent application demonstrated how chain-selective TUBEs can differentiate context-dependent ubiquitination of RIPK2: K63-TUBEs specifically captured L18-MDP-induced K63 ubiquitination, while K48-TUBEs selectively recognized PROTAC-induced K48 ubiquitination of the same protein [43]. This approach provides a high-throughput compatible method for investigating linkage-specific ubiquitination dynamics in physiological contexts.
Mass spectrometry-based quantification strategies have also proven invaluable for identifying and quantifying branched ubiquitin linkages. The AQUA (Absolute QUAntification) method enabled the discovery that K48-K63 branched linkages are abundant in mammalian cells and increase in response to IL-1β stimulation [84]. Combined with linkage-specific antibodies and advanced genetic approaches, these tools continue to expand our understanding of the ubiquitin code in immunity.
The Scientist's Toolkit: Key Research Reagents
Table 4: Essential Research Reagents for Studying Ubiquitin Signaling
| Research Tool | Specific Example | Application/Function | Experimental Use |
|---|---|---|---|
| Linkage-Specific TUBEs | K63-TUBE, K48-TUBE, M1-TUBE [43] | Selective enrichment of specific ubiquitin chain types | Capture endogenous ubiquitinated proteins for immunoblotting or proteomics |
| LUBAC Inhibitors | HOIP-targeting compounds [4] | Block linear ubiquitination | Determine LUBAC-specific functions in signaling pathways |
| DUB Inhibitors | OTULIN inhibitors, CYLD inhibitors [43] | Prevent specific deubiquitination | Probe functions of specific ubiquitin chain types |
| Linkage-Specific Antibodies | Anti-K63-Ub, Anti-K48-Ub, Anti-M1-Ub [83] | Detect specific ubiquitin linkages | Immunoblotting, immunofluorescence for chain type identification |
| Activity-Based Probes | Ub-VS, Ub-PA [38] | Label active DUBs and E enzymes | Identify active components of ubiquitination machinery |
| Mutant Ubiquitin Plasmids | Ub(K48R), Ub(K63R), Ub(K0) [83] | Define linkage requirements | Express in cells to determine chain type necessity for specific processes |
The functional divergence between M1, K63, and K48 ubiquitin chains represents a fundamental mechanism for achieving specificity and precision in innate immune signaling. While each chain type possesses characteristic functions—K48 in degradation, K63 in scaffolding, and M1 in inflammatory regulation—their interplay through sequential action, competition, and branching creates a sophisticated regulatory network that precisely controls immune responses. The recent discovery of STING-induced LUBAC activation and the protective function of K48-K63 branched chains against deubiquitination exemplify how continued research reveals unexpected complexities in ubiquitin-dependent signaling. Understanding these mechanisms not only advances fundamental knowledge of immune regulation but also unveils novel therapeutic opportunities for inflammatory diseases, autoimmune disorders, and cancer where ubiquitin pathway components are frequently dysregulated.
The ubiquitin code represents a sophisticated post-translational modification system where diverse ubiquitin chain architectures encode specific cellular signals. Among these, heterotypic M1/K63-branched ubiquitin chains have emerged as critical regulators of immune signaling pathways, particularly in nuclear factor kappa B (NF-κB) activation and inflammatory responses. These chains combine the signaling properties of linear M1-linked ubiquitin with K63-linked ubiquitin to create unique platforms for protein assembly and signal amplification. This technical review examines the current understanding of M1/K63-branched chain formation, structural characteristics, functional roles in immune regulation, and experimental approaches for their study. We highlight how these complex ubiquitin architectures facilitate crosstalk between different signaling pathways and discuss their implications for therapeutic intervention in inflammatory diseases and cancer.
Ubiquitination is a reversible post-translational modification that regulates virtually every cellular process in eukaryotes, with particular importance in immune system function. The versatility of ubiquitin signaling stems from its ability to form diverse chain architectures through eight different linkage types: seven via lysine residues (K6, K11, K27, K29, K33, K48, K63) and one via the N-terminal methionine (M1). These chains can be homotypic (single linkage type), mixed (multiple linkages in linear sequence), or branched (multiple linkages on a single ubiquitin molecule) [5] [69]. The specific topology of ubiquitin chains determines their recognition by effector proteins containing ubiquitin-binding domains (UBDs), thereby dictating functional outcomes.
Heterotypic branched ubiquitin chains represent an advanced layer of complexity in the ubiquitin code. These structures contain ubiquitin monomers modified at two or more acceptor sites simultaneously, creating branching points that can be recognized by specialized readers [69]. Branched chains account for approximately 10-20% of total ubiquitin polymers in cells, with K11/K48, K29/K48, K48/K63, and M1/K63 among the best-characterized types [86]. The formation of these complex architectures enables integration of signals from different ubiquitin-dependent pathways and can determine substrate fate with greater specificity than homotypic chains.
The focus of this review, M1/K63-branched ubiquitin chains, occupies a critical position at the intersection of inflammatory signaling and cell death pathways. These heterotypic chains combine the scaffold-forming properties of K63-linked ubiquitin with the unique signaling capabilities of linear M1-linked ubiquitin, creating a platform that preferentially recruits specific effector proteins to regulate immune responses [5] [2]. Understanding the formation, recognition, and function of these complex ubiquitin signals is essential for deciphering their roles in health and disease.
Molecular Machinery for M1/K63-Branched Ubiquitin Chain Formation
The Linear Ubiquitin Chain Assembly Complex (LUBAC)
Linear ubiquitination is exclusively catalyzed by the linear ubiquitin chain assembly complex (LUBAC), a ~600 kDa complex composed of three core subunits: HOIP (RNF31), HOIL-1L (RBCK1), and SHARPIN [5] [2]. HOIP serves as the catalytic center of LUBAC, containing a RING-between-RING (RBR) domain and a unique C-terminal linear ubiquitin chain determining domain (LDD) that specifically positions the N-terminus of acceptor ubiquitin for M1-linkage formation [5]. The catalytic activity of full-length HOIP is autoinhibited by its N-terminal domain, and this inhibition is relieved when HOIL-1L and SHARPIN bind to the ubiquitin-associated domain (UBA) of HOIP through their ubiquitin-like (UBL) domains [5].
HOIL-1L and SHARPIN play crucial structural and regulatory roles in LUBAC function. These subunits interact with each other via LUBAC-tethering motifs (LTMs) that fold into a single globular domain, stabilizing the trimeric complex [5]. HOIL-1L also exhibits catalytic activity as an atypical E3 ligase that can form oxyester bonds between ubiquitin and serine/threonine residues of substrates, potentially serving as a priming step for subsequent linear ubiquitination [5]. Additionally, HOIL-1L mediates monoubiquitination of all LUBAC subunits, which attenuates LUBAC function by providing substrates for HOIP-mediated linear ubiquitination [2].
Table 1: Core Components of the Linear Ubiquitin Chain Assembly Complex (LUBAC)
| Component | Gene | Molecular Weight | Domain Architecture | Function in LUBAC |
|---|---|---|---|---|
| HOIP | RNF31 | 58 kDa | NZF, UBA, RBR, LDD | Catalytic subunit; only E3 capable of forming M1-linked chains |
| HOIL-1L | RBCK1 | 123 kDa | UBL, RBR, NZF | Stabilizing subunit; regulates activity via monoubiquitination |
| SHARPIN | SHARPIN | 40 kDa | UBL, PH, NZF | Adaptor subunit; essential for complex stability and recruitment |
Collaborative E2 Enzymes and Priming Requirements
LUBAC functions with the ubiquitin-conjugating enzyme E2 L3 (UBE2L3) to transfer ubiquitin to substrates [2]. The assembly of linear ubiquitin chains by HOIP requires priming of the first ubiquitin on a substrate lysine residue, followed by linkage of an incoming ubiquitin to the N-terminus of the primed target ubiquitin [5]. Notably, HOIP demonstrates a preference for assembling linear ubiquitin chains on K63-ubiquitinated substrates, resulting in the formation of heterotypic M1/K63-branched ubiquitin chains [5]. This priming mechanism provides a direct structural basis for crosstalk between M1- and K63-linked ubiquitination pathways.
Several E3 ligases collaborate with LUBAC to facilitate M1/K63-branched chain formation. The RBR E3 ligase Parkin enhances LUBAC-mediated linear ubiquitination of NEMO by first modifying NEMO with K63-linked ubiquitin, creating a priming platform for subsequent linear chain extension [5]. Similarly, TRAF6, a RING E3 ligase involved in NF-κB activation, synthesizes K63-linked chains that serve as substrates for LUBAC-mediated branching [69]. This collaborative mechanism between E3s with different linkage specificities enables temporal and spatial control of branched chain formation in response to specific cellular signals.
Structural Basis of M1/K63-Branched Chain Formation
The structural organization of M1/K63-branched chains creates unique binding interfaces recognized by specific effector proteins. In these heterotypic structures, a single ubiquitin molecule serves as a branch point where both M1- and K63-linked chains converge. This architecture potentially allows simultaneous engagement of readers specific for each linkage type, facilitating the assembly of multiprotein signaling complexes [69]. The M1 linkage forms a rigid, extended structure that contrasts with the more flexible conformations of K63-linked chains, potentially creating a specific three-dimensional topology that enhances binding avidity for certain UBDs [5].
Table 2: Enzymatic Machinery Involved in M1/K63-Branched Ubiquitin Chain Formation
| Enzyme | Type | Linkage Specificity | Role in M1/K63 Branch Formation |
|---|---|---|---|
| UBE2L3 | E2 | M1-linear | Primary E2 for LUBAC-mediated linear ubiquitination |
| Ubc13/Uev1A | E2 | K63 | E2 complex for TRAF6-mediated K63 ubiquitination |
| HOIP | E3 (RBR) | M1-linear | Catalytic core of LUBAC; extends M1 chains on K63-primed substrates |
| TRAF6 | E3 (RING) | K63 | Synthesizes K63 chains that serve as primers for LUBAC |
| Parkin | E3 (RBR) | K63, K48, K11, K6 | Primes substrates with K63 chains for LUBAC recruitment |
Functional Roles in Immune Signaling Pathways
Regulation of NF-κB Signaling
M1/K63-branched ubiquitin chains play a pivotal role in the activation of NF-κB signaling pathways downstream of various immune receptors, including tumor necrosis factor receptor (TNFR), Toll-like receptors (TLRs), and interleukin-1 receptor (IL-1R) [75] [2]. Upon TNFR stimulation, LUBAC modifies components of the signaling complex with M1/K63-branched chains, particularly targeting the NF-κB essential modulator (NEMO) [2]. This modification creates a platform for recruitment and activation of the IκB kinase (IKK) complex, which phosphorylates the inhibitor of NF-κB (IκBα), leading to its K48-linked ubiquitination and proteasomal degradation [75].
The binding of NEMO to M1/K63-branched chains occurs through its UBAN (UBD in ABIN and NEMO) domain, which exhibits high affinity for linear ubiquitin via a surface on the proximal ubiquitin moiety and the canonical Ile44 surface on the distal ubiquitin [2]. This interaction promotes IKK oligomerization and activation, ultimately resulting in nuclear translocation of NF-κB and transcription of pro-inflammatory genes [2]. The presence of both M1 and K63 linkages in branched structures may enhance the stability of signaling complexes and increase the specificity of pathway activation compared to homotypic chains.
Modulation of Inflammasome Activation and Cell Death
Beyond NF-κB signaling, M1/K63-branched ubiquitin chains contribute to the regulation of inflammasome activation and cell death pathways. In sepsis and other inflammatory conditions, linear ubiquitination of receptor-interacting protein kinase 1 (RIPK1) by LUBAC, in conjunction with K63-linked ubiquitination, helps regulate the balance between pro-survival signaling and cell death induction [75]. The deubiquitinating enzyme USP5 removes K63-linked chains from RIPK1 to inhibit its activity, thereby suppressing the necroptosis pathway and protecting cardiomyocytes from septic injury [75].
The interplay between different ubiquitin linkage types in these pathways creates a sophisticated regulatory network that determines cellular fate in response to inflammatory stimuli. M1/K63-branched chains appear to favor pro-survival signaling, while their disassembly or modification can shift the balance toward inflammatory cell death. This delicate equilibrium is essential for maintaining appropriate immune responses and preventing excessive tissue damage during infection or sterile inflammation.
Experimental Approaches for Studying M1/K63-Branched Ubiquitin Chains
Detection and Validation Methodologies
The study of M1/K63-branched ubiquitin chains requires specialized methodologies due to their low abundance, transient nature, and structural complexity. Several experimental approaches have been developed to detect and validate these heterotypic structures:
Ubiquitin Binding Assays and Mass Spectrometry: Ubiquitin interactor pulldown coupled with mass spectrometry enables identification of proteins with binding preference for specific ubiquitin chain architectures [87]. This approach can elucidate branch-specific interactors by using immobilized ubiquitin chains of defined length and linkage composition as bait. Recent studies have employed K48- and K63-linked ubiquitin chains of varying lengths, along with heterotypic branched chains, to identify novel branch-specific binders [87].
Surface Plasmon Resonance (SPR): SPR provides quantitative analysis of binding affinity and specificity between branched ubiquitin chains and potential readers. This technique can distinguish binders with preference for branched chains over homotypic chains by comparing binding kinetics to different ubiquitin architectures [88]. For example, SPR has been used to validate the K48/K63 branch-specific binding of huntingtin-interacting protein HIP1 [87].
Linkage-Specific Antibodies: Bispecific antibodies that recognize two different linkage types simultaneously have been developed for the detection of heterotypic ubiquitin chains [88]. While challenging to generate, these reagents can function as "coincidence detectors" that gain avidity from simultaneous detection of multiple linkages, enabling specific recognition of branched chains over homotypic or mixed chains [88].
UbiCRest Assay: This method uses linkage-specific deubiquitinases (DUBs) to selectively disassemble ubiquitin chains, allowing determination of chain composition and complexity [87]. For M1/K63-branched chains, sequential treatment with K63-specific (AMSH) and M1-specific (OTULIN) DUBs can confirm the presence of both linkages within the same polymer.
Consideration of DUB Inhibitors in Experimental Design
The choice of deubiquitinase inhibitors during pulldown experiments significantly impacts the stability of ubiquitin chains and consequently affects the identification of interactors. Comparative studies using chloroacetamide (CAA) and N-ethylmaleimide (NEM) have revealed inhibitor-dependent variations in ubiquitin interactors [87]. NEM provides more complete inhibition of chain disassembly but has greater potential for off-target effects due to alkylation of exposed cysteines on non-DUB proteins, which could alter ubiquitin-binding surfaces [87]. In contrast, CAA is more cysteine-specific but allows partial chain disassembly during experiments [87]. Researchers must carefully consider these factors when designing studies of M1/K63-branched chains and interpreting resulting data.
Diagram 1: Experimental Workflow for M1/K63-Branched Ubiquitin Chain Interactor Identification
Research Reagent Solutions for M1/K63-Branched Ubiquitin Studies
Table 3: Essential Research Reagents for Studying M1/K63-Branched Ubiquitin Chains
| Reagent Category | Specific Examples | Function/Application | Considerations |
|---|---|---|---|
| E3 Ligase Complexes | LUBAC (HOIP/HOIL-1L/SHARPIN), TRAF6, Parkin | In vitro synthesis of M1/K63-branched chains | Co-expression systems may be needed for proper complex assembly |
| E2 Conjugating Enzymes | UBE2L3 (for M1), Ubc13/Uev1A (for K63) | Linkage-specific ubiquitin chain elongation | E2 specificity determines linkage type in reconstitution assays |
| Deubiquitinases | OTULIN (M1-specific), AMSH (K63-specific) | Chain validation, UbiCRest assays | Specificity must be verified for interpretation of cleavage patterns |
| Linkage-Specific Antibodies | K11/K48-bispecific, K48-specific, K63-specific, M1-specific | Detection, Western blotting, immunoprecipitation | Bispecific antibodies enable branched chain detection |
| DUB Inhibitors | Chloroacetamide (CAA), N-ethylmaleimide (NEM) | Stabilize ubiquitin chains during pulldowns | NEM more potent but less specific than CAA |
| Ubiquitin Mutants | K63R, K48R, M1-only (K0) | Control for linkage specificity, priming requirements | K0 ubiquitin (all lysines mutated) useful for exclusive M1 chain formation |
| Mass Spectrometry Standards | Ub-AQUA (Absolute QUAntification) peptides | Quantitative mass spectrometry | Isotope-labeled internal standards for precise quantification |
The study of M1/K63-branched ubiquitin chains represents a frontier in our understanding of how complex ubiquitin architectures encode sophisticated biological information. These heterotypic chains serve as critical integration points for multiple signaling pathways, particularly in the regulation of inflammatory responses and cell fate decisions. The collaborative synthesis of these chains by E3 ligases with different linkage specificities, such as LUBAC working in concert with TRAF6 or Parkin, provides a mechanism for signal amplification and specificity in immune signaling.
Future research directions should focus on developing more sensitive and specific tools for detecting endogenous M1/K63-branched chains, elucidating the structural basis of their recognition by reader proteins, and understanding how dysregulation of their formation or disassembly contributes to human diseases. The emerging role of M1/K63-branched chains in cancer pathogenesis [2] and sepsis [75] suggests these structures may represent valuable therapeutic targets for modulating immune responses in disease contexts. As our technical capabilities for studying these complex ubiquitin signals advance, so too will our understanding of their fundamental roles in cellular regulation and our ability to target them for therapeutic benefit.
Comparative Analysis of M1-Ubiquitin Roles in Macrophage Polarization vs. T-Cell Activation
M1-linked linear ubiquitin chains, assembled by the Linear Ubiquitin Chain Assembly Complex (LUBAC), represent a critical post-translational modification regulating immune cell signaling and function. This review provides a comparative analysis of M1-ubiquitin's roles in macrophage polarization and T-cell activation, highlighting its cell-type-specific mechanisms. In macrophages, M1-ubiquitination directly regulates polarization dynamics by controlling NF-κB signaling, inflammasome activation, and metabolic reprogramming. While M1-ubiquitin's functions in T-cell activation are less characterized, emerging evidence suggests important roles in regulating T-cell receptor signaling and effector functions. The striking contrast between well-defined macrophage pathways and emerging T-cell mechanisms underscores significant knowledge gaps and reveals compelling opportunities for therapeutic targeting of immune disorders through manipulation of linear ubiquitination pathways.
Linear ubiquitination, characterized by the covalent attachment of ubiquitin molecules through methionine-1 (M1) linkages, serves as a specialized signaling mechanism in immune regulation. Unlike conventional K48-linked chains that target proteins for proteasomal degradation, M1-linked ubiquitin chains function primarily as scaffolding elements that facilitate protein-protein interactions and activate key signaling pathways [63] [75]. The assembly of these chains is exclusively catalyzed by the Linear Ubiquitin Chain Assembly Complex (LUBAC), comprising HOIP, HOIL-1L, and SHARPIN subunits [63].
The M1-ubiquitin system operates as a precise regulatory mechanism in innate and adaptive immunity. In macrophages, M1-ubiquitination directly controls polarization states—the process by which macrophages adopt pro-inflammatory (M1-like) or anti-inflammatory (M2-like) phenotypes in response to environmental cues [32] [33]. This review systematically compares the established mechanisms of M1-ubiquitin in macrophage polarization against its emerging roles in T-cell activation, highlighting both conserved principles and cell-type-specific functions while identifying critical knowledge gaps in our current understanding of T-cell linear ubiquitination.
M1-Ubiquitin in Macrophage Polarization
Molecular Mechanisms of M1-Ubiquitination in Macrophage Signaling
Macrophage polarization is intricately regulated by M1-ubiquitination through several key signaling nodes. The NF-κB pathway represents a primary target, where LUBAC-generated M1 chains on NEMO (IKKγ) activate the IKK complex, driving transcription of pro-inflammatory genes characteristic of M1 polarization [75]. Recent research has identified that STING activation induces HOIP-mediated synthesis of M1 ubiquitin chains to stimulate NF-κB signaling, connecting DNA sensing to macrophage polarization [31]. This STING-HOIP axis promotes NF-κB and IRF3 activation in human THP1 monocytes and mouse bone marrow-derived macrophages independently of STING's role in LC3B lipidation [31].
The deubiquitinases OTULIN and CYLD provide counter-regulation by hydrolyzing M1-linked ubiquitin chains, preventing excessive inflammation. OTULIN deficiency causes ligand-independent NF-κB activation and severe inflammation, mimicking human OTULIN-related autoinflammatory syndrome (ORAS) [32] [33]. Similarly, CYLD removes K63-linked chains from NF-κB and JNK adaptors, with CYLD-deficient macrophages exhibiting hyperinflammatory signaling and increased susceptibility to colitis-associated colorectal cancer [32].
Table 1: Key E3 Ligases and Deubiquitinases Regulating Macrophage Polarization via M1-Ubiquitination
| Enzyme | Type | Target/Pathway | Effect on Polarization |
|---|---|---|---|
| LUBAC (HOIP/HOIL-1L/SHARPIN) | E3 Ligase Complex | NEMO/IKK complex, STING, TNFR | Promotes M1 polarization via NF-κB activation |
| OTULIN | Deubiquitinase | Hydrolyzes M1 chains on TLR/TNF adaptors | Restrains M1 polarization; prevents excessive inflammation |
| CYLD | Deubiquitinase | Removes K63/M1 chains from NF-κB/JNK adaptors | Limits M1 cytokine output; deficiency promotes hyperinflammation |
| A20 | Ubiquitin-editing Enzyme | TRAF6, RIP1, NEMO in NF-κB/MAPK pathways | Terminates M1 signaling; edits ubiquitin chains on key adaptors |
M1-Ubiquitin Regulation of Inflammatory Signaling Pathways
M1-ubiquitination regulates multiple inflammatory pathways that drive macrophage polarization. Upon TLR engagement, LUBAC synthesizes M1 chains that recruit the IKK complex through NEMO, leading to IκBα phosphorylation and degradation, and subsequent nuclear translocation of NF-κB [75]. This process enables transcription of M1-associated genes including TNF-α, IL-6, IL-1β, and iNOS [32] [33].
Beyond NF-κB signaling, M1-ubiquitination regulates inflammasome assembly and activity. The deubiquitinase BRCC3, as part of the BRISC complex, removes K48/K63 ubiquitin from NLRP3, permitting ASC oligomerization, caspase-1 activation, and IL-1β maturation [32]. This deubiquitination step represents a critical checkpoint for inflammasome activation, with BRCC3 inhibition blocking IL-1β release [32] [33].
Table 2: Quantitative Effects of M1-Ubiquitination on Macrophage Polarization Markers
| Experimental Manipulation | M1 Marker Expression | M2 Marker Expression | Functional Outcome |
|---|---|---|---|
| HOIP deletion (in BMDMs) | Reduced TNF-α, IL-6, IL-1β | Not reported | Impaired bacterial clearance; reduced inflammation |
| OTULIN deficiency | Increased TNF-α, IL-6, IL-1β | Not reported | Severe autoinflammation; ORAS pathology |
| CYLD knockout | Enhanced TNF-α, IL-6 production | Not reported | Hyperinflammatory signaling; increased colitis-associated cancer |
| STING-induced HOIP activation | Enhanced NF-κB target genes | Not reported | Augmented pro-inflammatory response to cytosolic DNA |
Experimental Approaches for Studying M1-Ubiquitin in Macrophages
Current methodologies for investigating M1-ubiquitination in macrophage polarization encompass molecular, biochemical, and cellular approaches:
Polarization Protocols: Bone marrow-derived macrophages (BMDMs) are generated by culturing bone marrow cells with M-CSF (20 ng/mL) for 5-7 days [89]. For M1 polarization, cells are stimulated with LPS (100 ng/mL) and IFN-γ (20 ng/mL) for 24 hours; for M2 polarization, IL-4 (20 ng/mL) and IL-13 (20 ng/mL) are used [89].
Ubiquitination Assays: Co-immunoprecipitation followed by western blotting with M1-linkage specific antibodies (e.g., anti-linear ubiquitin specific antibodies) detects M1-ubiquitination. Overexpression of LUBAC components or OTULIN/CYLD mutants modulates chain formation [31].
Functional Measurements: Cytokine secretion profiles (TNF-α, IL-6, IL-10) are quantified using multiplex immunoassays or ELISA. Metabolic parameters including glycolysis and oxidative phosphorylation are assessed via Seahorse analyzers [32].
M1-Ubiquitin in T-Cell Activation
Current Understanding of M1-Ubiquitin in T-Cell Biology
While the role of M1-ubiquitination in macrophage polarization is well-established, its functions in T-cell activation remain less characterized. Current evidence suggests that LUBAC-mediated linear ubiquitination regulates T-cell receptor (TCR) signaling and subsequent immune responses. In T-cells, M1-ubiquitin chains likely modulate key signaling nodes downstream of TCR engagement, potentially affecting activation thresholds, differentiation programs, and effector functions.
The available literature indicates that M1-ubiquitination may influence T-cell biology through regulation of NF-κB signaling, similar to mechanisms observed in macrophages. However, T-cell-specific functions likely involve unique substrates and regulatory mechanisms that remain to be fully elucidated. This knowledge gap presents significant opportunities for future research into cell-type-specific functions of linear ubiquitination in adaptive immunity.
Comparative Signaling Mechanisms
Despite limited T-cell-specific data, comparative analysis reveals that M1-ubiquitination likely regulates conserved signaling nodes across immune cell types while also exhibiting cell-type-specific functions:
NF-κB Pathway: Both macrophages and T-cells utilize M1-ubiquitination for optimal NF-κB activation, though likely through distinct receptor systems (TLRs/TNFR in macrophages vs. TCR in T-cells).
Cell Death Regulation: M1-ubiquitination regulates necroptosis sensitivity in macrophages, with M1-polarized macrophages showing enhanced susceptibility to RIPK3/MLKL-dependent necroptosis [90]. Similar mechanisms may operate in T-cells to control activation-induced cell death.
Metabolic Reprogramming: Emerging evidence suggests M1-ubiquitination integrates immune signaling with metabolic pathways in macrophages [32] [33], potentially paralleling metabolic changes during T-cell activation.
Research Reagent Solutions
Table 3: Essential Research Reagents for Studying M1-Ubiquitin in Immune Cells
| Reagent/Category | Specific Examples | Research Application | Experimental Function |
|---|---|---|---|
| Polarization Inducers | LPS, IFN-γ, IL-4, IL-13 | Macrophage polarization | Induce M1 (LPS+IFN-γ) or M2 (IL-4+IL-13) phenotypes |
| Cytokine Detection | Bio-Plex Pro Mouse Cytokine 8-Plex assay, ELISA kits | Cytokine profiling | Quantify secreted cytokines (TNF-α, IL-6, IL-10, etc.) |
| Ubiquitination Modulators | LUBAC expression plasmids, OTULIN/CYLD mutants, HOIP inhibitors | Ubiquitination studies | Modulate M1-ubiquitin chain formation or removal |
| Cell Culture Models | Bone marrow-derived macrophages, Peritoneal macrophages, THP-1 cells | In vitro studies | Provide biologically relevant systems for polarization assays |
| Signaling Inhibitors | RIPK3 inhibitors (GSK872, GSK843), IKK inhibitors, Proteasome inhibitors | Pathway analysis | Dissect specific signaling contributions to polarization |
| Antibodies | Anti-linear ubiquitin, Anti-NEMO, Anti-HOIP, Anti-OTULIN | Detection and quantification | Identify protein expression and ubiquitination status |
This comparative analysis reveals the well-established role of M1-ubiquitination in macrophage polarization while highlighting significant knowledge gaps regarding its functions in T-cell activation. In macrophages, LUBAC-generated M1 chains directly regulate polarization states through precise control of NF-κB signaling, inflammasome activation, and metabolic reprogramming, with deubiquitinases like OTULIN and CYLD providing critical counter-regulation.
Future research should prioritize elucidating T-cell-specific functions of M1-ubiquitination, particularly its roles in TCR signaling, activation-induced cell death, and T-cell differentiation. The development of cell-type-specific knockout models and more sensitive tools for detecting endogenous M1-ubiquitination will be essential for advancing this field. Additionally, comparative studies examining M1-ubiquitination across different T-cell subsets (CD4+ vs. CD8+, naive vs. memory) may reveal subset-specific regulatory mechanisms.
From a therapeutic perspective, targeting M1-ubiquitination pathways holds significant promise for immune-mediated diseases. However, successful translation will require a more comprehensive understanding of cell-type-specific functions to minimize off-target effects. The striking contrast between well-defined macrophage pathways and emerging T-cell mechanisms underscores the complexity of immune regulation through linear ubiquitination and reveals compelling opportunities for future investigation.
Visualized Signaling Pathways
Benchmarking M1-Ubiquitin-Dependent NF-κB Activation Against Other Inflammatory Pathways
The nuclear factor-kappa B (NF-κB) pathway serves as a pivotal mediator of inflammatory responses, with its activation mechanisms fine-tuned by various post-translational modifications. Among these, M1-linked linear ubiquitination, catalyzed by the Linear Ubiquitin Chain Assembly Complex (LUBAC), has emerged as a crucial, specialized regulatory layer. This whitepaper provides a technical benchmark of M1-ubiquitin-dependent NF-κB activation against other inflammatory signaling pathways. We synthesize current mechanistic insights, quantitative dynamic data, and experimental methodologies, framing this analysis within the broader thesis that M1-linear ubiquitin chains constitute a master regulatory switch that integrates signaling from multiple innate immune receptors and cellular stress sensors. The content is structured to serve researchers, scientists, and drug development professionals seeking to understand and manipulate this specific arm of inflammatory signaling for therapeutic purposes.
NF-κB transcription factors regulate hundreds of genes involved in immunity, inflammation, and cell survival [21]. The canonical NF-κB activation pathway involves phosphorylation-induced degradation of IκB inhibitors, liberating primarily p50/RelA dimers for nuclear translocation [21]. A critical regulatory step in this process involves protein ubiquitination, which occurs through diverse chain linkage types that determine functional outcomes.
M1-linked ubiquitination represents a specialized mechanism wherein ubiquitin monomers form chains through sequential conjugation to the N-terminal methionine residue [18]. This linear ubiquitination is exclusively catalyzed by LUBAC, a multi-subunit E3 ubiquitin ligase complex composed of HOIP, HOIL-1L, and SHARPIN in mammals, with LUBEL serving as the functional homolog in Drosophila [18] [63]. Recent research has established that M1-ubiquitin chains do not typically target substrates for proteasomal degradation but instead function as scaffolding platforms that recruit signaling complexes through ubiquitin-binding domains, facilitating activation of the IKK complex and subsequent NF-κB signaling [18] [63].
The positioning of M1-ubiquitination within inflammatory signaling networks reveals its function as an integrative node that transduces signals from diverse stimuli into coordinated NF-κB activation. This review systematically benchmarks M1-ubiquitin-dependent NF-κB activation against other inflammatory pathways through quantitative dynamic modeling, mechanistic dissection, and experimental methodology comparison.
Mechanistic Foundations of M1-Ubiquitin Signaling
Structural and Biochemical Basis of Linear Ubiquitination
The catalytic activity for M1-ubiquitin chain formation resides primarily in the RBR (RING-In-Between-RING) domain and the adjacent Linear ubiquitin chain-Determining Domain (LDD) of HOIP, the catalytic core of LUBAC [18]. This unique structural arrangement positions two ubiquitin moieties in precise orientation for Met1 linkage formation, a mechanism distinct from lysine-linked ubiquitination [18]. Specific deubiquitinases provide negative regulation, with OTULIN hydrolyzing M1-linked chains and CYLD removing K63-linked and M1-linked chains [33] [63].
Table 1: Core Components of the M1-Ubiquitination Machinery
| Component | Function | Functional Characteristics |
|---|---|---|
| HOIP | Catalytic core of LUBAC | RBR-LDD domain confers M1-specific chain elongation capability |
| HOIL-1L | LUBAC accessory subunit | Stabilizes complex, regulates HOIP activity |
| SHARPIN | LUBAC accessory subunit | Essential for proper complex assembly and function |
| LUBEL | Drosophila HOIP homolog | Catalyzes M1-Ub chain formation in flies |
| OTULIN | Deubiquitinase | Specifically hydrolyzes M1-linked ubiquitin chains |
| CYLD | Deubiquitinase | Removes K63-linked and M1-linked chains |
Signaling Topology: M1-Ubiquitin in Pathway Architecture
The following diagram illustrates the position of M1-ubiquitin signaling within the broader NF-κB activation network, highlighting its integrative function across multiple pathway inputs:
Diagram 1: M1-Ubiquitin-Dependent NF-κB Signaling Network. M1-ubiquitination serves as a convergence point for signals from immune receptors, cytosolic sensors, and cellular stress pathways. The diagram highlights the central positioning of LUBAC-synthesized M1-linked ubiquitin chains as scaffolds that facilitate TAK1 and IKK complex activation, with key negative feedback mechanisms provided by deubiquitinases (OTULIN, CYLD) and the ubiquitin editor A20.
Quantitative Benchmarking of Pathway Dynamics
Temporal Activation Profiles
The dynamic characteristics of NF-κB activation differ substantially depending on the initiating stimulus and the involvement of M1-ubiquitination. Quantitative measurements in microglial cells reveal that TNFα stimulation induces biphasic NF-κB activation with an initial peak at approximately 20 minutes, followed by a secondary smaller amplitude peak around 90 minutes [91]. Upstream IKK activation is notably transient, peaking at 5 minutes and declining sharply to below 50% of maximal activity by 10 minutes post-stimulation [91].
Mathematical modeling of these dynamics suggests that the ubiquitin-proteasome system plays a more prominent role in regulating NF-κB activation kinetics than previously appreciated, with intermediate steps in the IKK-induced IκBα degradation pathway significantly influencing the temporal response profile [91]. This refined model required incorporation of nonlinear kinetics in IKK activation and inactivation to properly characterize the transient IKK activity observed experimentally.
Table 2: Kinetic Parameters of NF-κB Pathway Activation
| Stimulus | IKK Peak Activity | NF-κB Peak Activity | Oscillatory Behavior | M1-Ubiquitin Involvement |
|---|---|---|---|---|
| TNFα | 5 minutes | 20 minutes (primary) | Biphasic oscillation | Yes (LUBAC-dependent) |
| Lysosomal Damage | 15-30 minutes | 1-2 hours | Sustained activation | Yes (LUBAC/TAB-TAK1) |
| STING Activation | 1-2 hours | 2-4 hours | Monophasic sustained | Yes (HOIP-mediated) |
| IL-1/TLR | 5-15 minutes | 20-30 minutes | Biphasic oscillation | Yes (LUBAC-dependent) |
| Cellular Stress | Variable (10-30 min) | 30-90 minutes | Context-dependent | Yes (LUBEL/LUBAC) |
Signal Amplification and Threshold Properties
M1-ubiquitin-dependent NF-κB activation demonstrates distinctive signal amplification properties. Research in Drosophila reveals that LUBEL-mediated M1-ubiquitination creates a threshold response to hypoxic, oxidative, and mechanical stress, with flies exhibiting significantly reduced survival under these conditions when LUBEL function is compromised [18]. This protective function requires multiple components of the Immune Deficiency (Imd) pathway, including Tak1, Diap2, Kenny, and Dredd, but interestingly does not require the membrane receptor PGRP-LC, indicating a specific role in sterile inflammation rather than pathogen response [18].
In mammalian systems, LUBAC-mediated M1-ubiquitination provides signal amplification at multiple levels. At damaged lysosomes, M1-ubiquitin chains create localized signaling platforms that facilitate TAK1-dependent NF-κB activation, modulating gene expression for cell survival [92]. Similarly, during STING signaling, HOIP-mediated synthesis of M1 ubiquitin chains at Golgi-related membranes stimulates both NF-κB and IRF3 signaling branches, despite not being required for STING activation per se [31].
Experimental Benchmarking Methodologies
Standardized Reporter Assay Systems
Robust quantification of NF-κB pathway activation requires carefully validated reporter systems. The HEK293 NF-κB GFP-luciferase reporter cell line (TR860A-1 from System Biosciences) provides a well-characterized platform for benchmarking agonists and antagonists [93] [94]. Standardized protocols using this system include:
- Cell culture: Maintenance in DMEM with 10% FBS and 1% penicillin/streptomycin at 37°C and 5% CO₂
- Stimulation parameters: 5 ng/ml TNFα for 24 hours as optimal activation conditions
- Readout measurement: Luciferase reporter assays (Promega) with luminescence quantification
- Parallel viability assessment: CellTiter-Glo 2.0 assays to control for cytotoxicity
This standardized approach enables quantitative comparison of NF-κB activation across different stimuli and inhibitor treatments, with EC₅₀ values calculable using GraphPad Prism software [93].
M1-Ubiquitin Specific Detection Methods
Specialized methodologies have been developed to specifically monitor M1-ubiquitination events:
- M1-TUBE (Tandem Ubiquitin-Binding Entity): GST-tagged recombinant M1-specific TUBE domains for isolation of M1-ubiquitin chains from cell lysates [18]
- LUBAC inhibition: Mutagenesis or pharmacological inhibition of the RBR domain of HOIP to specifically block M1-ubiquitin chain formation [18]
- Deubiquitinase specificity profiling: Using catalytically active OTULIN versus other DUBs to verify M1-linkage specificity [33]
The experimental workflow below illustrates a comprehensive approach for benchmarking M1-ubiquitin-dependent NF-κB activation:
Diagram 2: Experimental Workflow for Benchmarking M1-Ubiquitin-Dependent NF-κB Activation. The diagram outlines key methodological approaches for quantifying M1-ubiquitin-specific contributions to NF-κB signaling, incorporating specific detection methods, inhibition strategies, and computational analysis approaches.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Studying M1-Ubiquitin-Dependent NF-κB Activation
| Reagent/Catalog Number | Type | Specific Function | Application Context |
|---|---|---|---|
| HEK293 NF-κB GFP-luciferase (TR860A-1) | Reporter cell line | Quantitative NF-κB activity monitoring | Pathway agonist/antagonist screening [93] |
| M1-TUBE | Recombinant binding protein | Selective isolation of M1-linked ubiquitin chains | Detection of M1-ubiquitination in cell lysates [18] |
| Recombinant TNFα (315-01A) | Cytokine | Canonical NF-κB pathway activation | Positive control for NF-κB stimulation [93] |
| HOIP/HOIL-1/SHARPIN antibodies | Immunological reagents | Detection of LUBAC components | Western blot, immunoprecipitation [18] |
| OTULIN expression constructs | Molecular biology reagent | Specific hydrolysis of M1-linked ubiquitin chains | Mechanistic studies of M1-chain function [33] |
| IKK-16 (S2882) | Small molecule inhibitor | IKKβ inhibition (IC₅₀ = 40 nM) | Benchmarking against upstream ubiquitin-dependent activation [93] |
| Bay 11-7082 (S2913) | Small molecule inhibitor | IκBα phosphorylation inhibition | Negative control for NF-κB blockade [93] |
Comparative Pathway Architecture
M1-Ubiquitin Versus Other Ubiquitin Linkages in NF-κB Activation
The NF-κB pathway integrates signals from multiple ubiquitin linkage types, each with distinct functional consequences:
K63-linked ubiquitination: Primarily involved in receptor-proximal signaling, facilitating recruitment of TAK1 and IKK complexes through adaptor proteins like RIP1. Serves as an activation signal without targeting substrates for degradation [92].
M1-linked ubiquitination: Functions as a scaffolding platform at multiple signaling nodes, including TNFR1, IL-1R, and TLR complexes. Amplifies signals and facilitates sustained NF-κB activation through LUBAC-mediated assembly of linear ubiquitin chains on substrates like NEMO [18] [63].
K48-linked ubiquitination: Principally targets proteins for proteasomal degradation, including IκBα and some negative regulators of NF-κB signaling. Creates negative feedback loops that terminate signaling [91].
Recent research reveals sophisticated crosstalk mechanisms between different ubiquitin linkage types. For instance, during lysosomal damage response, K63-linked ubiquitin chains initially recruit TAB2/3, which then facilitates LUBAC-mediated M1-ubiquitin synthesis and subsequent NF-κB activation [92]. This hierarchical ubiquitination creates a temporally regulated signaling cascade that ensures appropriate cellular responses to endolysosomal damage.
Context-Specific Pathway Utilization
The relative importance of M1-ubiquitin-dependent signaling varies significantly across biological contexts:
Sterile Inflammation: During hypoxic, oxidative, or mechanical stress, M1-ubiquitination through LUBEL (in Drosophila) or LUBAC (in mammals) becomes essential for NF-κB activation and survival, operating independently of pathogen recognition receptors [18].
STING Signaling: HOIP-mediated M1-ubiquitin chain synthesis occurs at Golgi-related membranes following STING activation, contributing to both NF-κB and IRF3 signaling without being required for initial STING activation [31].
Macrophage Polarization: M1-like pro-inflammatory macrophage polarization involves LUBAC-mediated linear ubiquitination of multiple signaling intermediates, creating a positive feedback loop that sustains the inflammatory phenotype [33].
Lysosomal Damage Response: Damaged lysosomes recruit LUBAC components that synthesize M1-ubiquitin chains, facilitating local NF-κB activation that promotes cell survival through expression of anti-apoptotic genes [92] [63].
Therapeutic Implications and Research Applications
The strategic positioning of M1-ubiquitination within inflammatory signaling networks makes it an attractive target for therapeutic intervention. Several approaches show promise:
LUBAC-specific inhibitors: Compounds targeting the RBR domain of HOIP to selectively block M1-ubiquitin chain formation without affecting other ubiquitination pathways [18].
OTULIN-based strategies: Modulation of M1-ubiquitin chain stability through regulation of OTULIN activity or expression [33].
Context-specific pathway manipulation: Selective targeting of M1-ubiquitin signaling in specific pathological contexts, such as chronic inflammatory diseases or cancer, where this pathway is dysregulated [63].
For research applications, benchmarking M1-ubiquitin-dependent NF-κB activation provides critical insights for:
- Target validation studies: Establishing the relative contribution of M1-ubiquitination to specific disease processes
- Drug screening platforms: Developing specialized assays that specifically monitor M1-ubiquitin-dependent signaling
- Combination therapy strategies: Identifying synergistic interactions between M1-ubiquitination inhibitors and other targeted therapies
- Biomarker development: Discovering signatures of M1-ubiquitin pathway activation in clinical samples
M1-ubiquitin-dependent NF-κB activation represents a specialized signaling mechanism that distinguishes itself from other inflammatory pathways through its unique positioning as an integrative signaling node, its scaffolding rather than degradative function, and its context-specific essentiality in sterile inflammation, organelle stress responses, and specific immune signaling branches. The benchmarking data presented in this technical guide provides researchers with quantitative parameters, standardized methodologies, and conceptual frameworks for investigating this critical regulatory pathway.
Future research directions should focus on developing more specific tools for manipulating M1-ubiquitination in physiological contexts, understanding the spatial regulation of LUBAC activity within cellular compartments, and translating these mechanistic insights into targeted therapeutic approaches for inflammatory diseases, cancer, and immune disorders. The continued benchmarking of M1-ubiquitin-dependent signaling against parallel inflammatory pathways will remain essential for understanding its unique contributions to immune homeostasis and disease pathogenesis.
The linear ubiquitin chain assembly complex (LUBAC) is a critical regulator of inflammatory signaling and innate immunity across metazoans. This whitepaper examines the deep evolutionary conservation between the Drosophila melanogaster E3 ligase LUBEL and its mammalian counterpart, LUBAC. We synthesize recent structural, functional, and mechanistic insights that position LUBEL as a powerful model system for deciphering the core principles of M1-linked ubiquitination in immune regulation. The conserved molecular architecture, shared substrate targeting strategies, and parallel signaling outputs reveal fundamental biological principles while identifying species-specific adaptations. For researchers and drug development professionals, this evolutionary perspective provides valuable frameworks for interrogating LUBAC function in human health and disease, offering novel methodological approaches and potential therapeutic strategies targeting the linear ubiquitination system.
Linear (M1-linked) ubiquitination represents a specialized post-translational modification where the C-terminal glycine of one ubiquitin molecule conjugates to the N-terminal methionine of another, forming head-to-tail polymeric chains. In mammals, this process is exclusively catalyzed by the multi-subunit linear ubiquitin chain assembly complex (LUBAC), comprising HOIP, HOIL-1L, and SHARPIN [4]. HOIP serves as the catalytic core containing the RING-IBR-RING (RBR) domain that confers M1-linkage specificity, while HOIL-1L and SHARPIN function as essential accessory subunits that stabilize the complex [95] [4].
The evolutionary roots of this system extend deeply into the animal kingdom. Drosophila melanogaster possesses a single homologous E3 ligase termed LUBEL (linear ubiquitin E3 ligase), which similarly catalyzes M1-linked ubiquitin chain formation [25]. Despite the simpler architecture—functioning without identified HOIL-1L or SHARPIN orthologs—LUBEL executes remarkably conserved immunological functions in fly immunity, establishing Drosophila as a potent genetic model for deciphering LUBAC biology.
This technical guide explores the fundamental conservation principles bridging Drosophila LUBEL and mammalian LUBAC, with emphasis on experimental approaches, signaling mechanisms, and research methodologies relevant to investigators studying M1-linked ubiquitination in immune signaling pathways.
Structural and Functional Conservation
Molecular Architecture and Catalytic Mechanisms
The core catalytic conservation between LUBEL and HOIP centers on the RBR domain architecture, which defines the RBR family of E3 ubiquitin ligases.
Table 1: Comparative Analysis of LUBEL and LUBAC Components
| Feature | Drosophila LUBEL | Mammalian LUBAC | Functional Significance |
|---|---|---|---|
| Catalytic Subunit | LUBEL (multiple isoforms) | HOIP (RNF31) | Both contain conserved RING1-IBR-RING2 domains with catalytic cysteine residue in RING2 for linear chain formation |
| Accessory Subunits | Not identified | HOIL-1L, SHARPIN | Mammalian complex stabilization; LUBEL may function as monomer or with unidentified partners |
| Domain Organization | RBR domain with ZnF, UBA, and LDD regions | RBR domain with ZnF, UBA, LDD, and PUB regions | Conservation of linear ubiquitin determining domain (LDD) essential for M1-specificity |
| Regulatory Mechanisms | Transcriptional regulation, potential protein partnerships | Neddylation, phosphorylation, OTULIN/CYLD binding | Differential complexity in regulatory inputs |
| Deubiquitinases | Not fully characterized | OTULIN, CYLD | Conservation of M1-chain hydrolysis machinery |
The catalytic mechanism involves a two-step transfer process: the RING1 domain recruits ubiquitin-charged E2 enzymes, followed by transthiolation of ubiquitin to a conserved cysteine residue in the RING2 domain, and finally iso-peptide bond formation with the N-terminal methionine of an acceptor ubiquitin [25] [4]. Both LUBEL and HOIP contain the linear ubiquitin determining domain (LDD) that recognizes acceptor ubiquitin and confers strict M1-linkage specificity.
Immune Signaling Pathway Conservation
Both LUBEL and LUBAC integrate into cognate immune signaling pathways with striking functional parallels, particularly in the regulation of NF-κB transcription factors.
Diagram 1: Conserved immune signaling pathways in Drosophila and mammals. LUBEL/LUBAC integration into NF-κB activation shows parallel architecture despite species-specific components.
The Drosophila LUBEL regulates the Imd pathway, which responds to Gram-negative bacterial infection through pattern recognition receptors of the PGRP family [25]. Upon immune activation, LUBEL synthesizes M1-linked chains on Kenny, the Drosophila IKKγ homolog, mirroring LUBAC-mediated modification of NEMO in mammalian NF-κB signaling [25]. This conservation extends to shrimp (Penaeus vannamei), where LUBEL homolog PvLUBEL mediates linear ubiquitination of hemocyanin, modulating immune responses to Vibrio parahaemolyticus and white spot syndrome virus [95].
Recent research reveals that mammalian LUBAC recruitment to specific cellular compartments is essential for signaling specificity. STING activation induces HOIP-mediated M1-ubiquitin chain synthesis at Golgi-related membranes, stimulating both NF-κB and IRF3 signaling [31]. This compartmentalization mirrors the observed tissue-specific regulation of LUBEL activity in Drosophila and shrimp immune tissues.
Experimental Approaches and Methodologies
Detection and Validation of Linear Ubiquitination
The identification and verification of M1-linked ubiquitin chains requires specialized methodologies due to the unique structural and biochemical properties of these modifications.
Table 2: Key Research Reagents for Linear Ubiquitination Studies
| Reagent/Category | Specific Examples | Application and Function | Considerations |
|---|---|---|---|
| Affinity Reagents | GST-NEMO-UBAN, chain-specific TUBEs | Selective enrichment of M1-linked chains from complex lysates | K63-TUBEs do not bind M1-chains, enabling linkage specificity |
| Detection Antibodies | Anti-linear ubiquitin specific antibodies | Immunoblotting, immunohistochemistry for M1-chain visualization | Specificity must be validated using OTULIN treatment controls |
| Deubiquitinases | Recombinant OTULIN, vOTU, CYLD | Linkage verification through selective chain cleavage | OTULIN specifically hydrolyzes M1-linkages; vOTU cleaves all except M1 |
| E3 Ligase Inhibitors | HOIPIN-1 | Specific covalent inhibition of HOIP/LUBEL catalytic activity | Useful for functional validation in cellular and in vivo models |
| Genetic Tools | RNAi, Minos transposable elements, CRISPR/Cas9 | Gene disruption, knockdown, and knockout studies | lubelMi flies show abolished infection-induced M1-chain formation |
| Infection Models | Ecc15, V. parahaemolyticus, WSSV, L18-MDP | Immune pathway stimulation in different species | Tissue-specific and pathogen-specific responses observed |
Protocol 1: Detection of M1-Linked Ubiquitin Chains Using UBAN Domain Affinity Purification
Cell Lysis: Prepare tissue or cell lysates in modified RIPA buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 1 mM EDTA) supplemented with 10 mM N-ethylmaleimide to preserve ubiquitin chains.
Affinity Purification: Incubate 500 μg of protein lysate with 20 μg of GST-tagged NEMO UBAN domain (or equivalent M1-chain binding module) pre-coupled to glutathione-sepharose beads for 2 hours at 4°C with rotation.
Washing: Pellet beads and wash three times with ice-cold lysis buffer containing 300 mM NaCl to reduce non-specific interactions.
Elution and Analysis: Elute bound proteins with 2× Laemmli buffer, resolve by SDS-PAGE, and transfer to PVDF membrane for immunoblotting with anti-ubiquitin antibodies.
Specificity Controls: Treat parallel samples with recombinant OTULIN (1 μg/reaction) for 1 hour at 37°C prior to analysis to verify M1-linkage specificity, as OTULIN selectively cleaves linear ubiquitin chains [25].
Protocol 2: Functional Validation Using Genetic Manipulation in Drosophila
Model Selection: Utilize available LUBEL mutant lines such as lubelMi (yw;Mi{ET1}LUBELMB00197) containing a 7.5 kb Minos transposable element insertion disrupting the catalytic RBR domain [25].
Immune Challenge: For oral infection studies, starve adult flies for 2 hours followed by feeding with Ecc15 Gram-negative bacteria (5×108 CFU/mL) in 5% sucrose solution for 16-24 hours.
Phenotypic Analysis:
- Survival Assay: Monitor and record mortality daily post-infection.
- Pathogen Clearance: Homogenize flies in sterile PBS and plate serial dilutions on LB agar to quantify bacterial loads.
- Gene Expression: Extract total RNA from whole flies or dissected tissues, followed by RT-qPCR analysis of antimicrobial peptide genes (Diptericin, Attacin) as Relish pathway readouts.
Biochemical Confirmation: Verify absence of infection-induced M1-ubiquitin chains in mutant flies using UBAN domain pull-down as described in Protocol 1.
Advanced Methodologies for Linkage-Specific Analysis
Recent technological advances enable more precise dissection of complex ubiquitination patterns. Tandem ubiquitin binding entities (TUBEs) with linkage specificity provide a powerful high-throughput compatible platform for studying endogenous protein ubiquitination [42]. The application of chain-selective TUBEs in 96-well plate formats allows quantitative assessment of context-dependent ubiquitination dynamics, as demonstrated for RIPK2 in inflammatory signaling.
For investigating more complex ubiquitin architectures, including branched chains containing M1-linkages, tandem ubiquitin mass spectrometry approaches combined with linkage-specific deubiquitinases provide unprecedented resolution [69]. These methodologies reveal the collaborative synthesis of branched ubiquitin chains by E3 ligases with different linkage specificities, expanding the complexity of ubiquitin codes in immune regulation.
Evolutionary Adaptations and Divergence
While the core catalytic function remains conserved, LUBEL and LUBAC exhibit significant functional specializations reflecting their respective immunological niches.
Tissue and Immune Specificity: Drosophila LUBEL demonstrates compartmentalized functionality, being essential for immune responses to oral infection but dispensable for systemic immunity following septic injury [25]. This contrasts with the broader requirement for LUBAC across multiple mammalian immune cell types and signaling pathways.
Complexity of Regulation: Mammalian LUBAC undergoes sophisticated multilayered regulation including neddylation, phosphorylation, and competitive binding of deubiquitinases OTULIN and CYLD [4]. The regulatory mechanisms controlling LUBEL activity remain less characterized but appear equally crucial for signaling fidelity.
Pathogen Countermeasures: The differential regulation of LUBEL-mediated signaling by distinct pathogens in shrimp illustrates evolutionary adaptation at the host-pathogen interface [95]. Vibrio parahaemolyticus suppresses PvLUBEL expression and PvHMC linear ubiquitination, whereas WSSV enhances these parameters, suggesting pathogen-specific evasion strategies.
Diagram 2: Evolutionary conservation and divergence between Drosophila LUBEL and mammalian LUBAC. Core catalytic function is preserved while regulatory complexity and immunological roles have diversified.
Research Applications and Therapeutic Implications
The evolutionary insights from comparative LUBEL-LUBAC studies provide tangible research applications and therapeutic development opportunities.
Experimental Model Selection Guidelines
- Drosophila melanogaster: Optimal for genetic screening of conserved linear ubiquitination functions, particularly in epithelial immunity and host-pathogen interactions in the gastrointestinal tract [25].
- Mammalian Cell Systems: Essential for pathway biochemistry, proteomic profiling, and therapeutic compound validation, especially with THP-1 monocytes and bone marrow-derived macrophages [31].
- Crustacean Models: Emerging systems for studying LUBEL function in antimicrobial defense, particularly relevant for understanding economic impacts in aquaculture [95].
Therapeutic Target Considerations
The deep conservation of linear ubiquitination mechanisms validates this pathway as a high-value therapeutic target with applications across multiple disease contexts:
- Inflammatory Diseases: Inhibition of LUBAC assembly or catalytic activity represents a promising strategy for autoimmune and autoinflammatory conditions, with HOIPIN-1 providing a proof-of-concept for covalent LUBAC inhibition [95].
- Cancer Therapeutics: Modulating LUBAC-mediated cell survival signaling may enhance efficacy of conventional chemotherapeutics, particularly in hematological malignancies.
- Infection Management: Pathogen-specific manipulation of linear ubiquitination pathways offers novel antimicrobial strategies, as evidenced by divergent pathogen responses in shrimp models [95].
The evolutionary journey from Drosophila LUBEL to mammalian LUBAC reveals both remarkable conservation of core mechanism and informative divergence in regulatory complexity. The conserved RBR catalytic core with M1-linkage specificity represents a fundamental immunological module that has been maintained across approximately 600 million years of evolution, while accessory subunits and regulatory mechanisms have diversified to meet species-specific immunological demands.
For researchers and drug development professionals, this evolutionary perspective provides valuable insights for experimental design, model system selection, and therapeutic targeting. The methodological approaches and conceptual frameworks derived from comparative studies will continue to accelerate our understanding of linear ubiquitination in health and disease, ultimately informing novel therapeutic strategies for inflammatory disorders, cancer, and infectious diseases.
Conclusion
M1-linked ubiquitination is established as a master regulator of inflammatory and cell survival signaling, with its precise manipulation holding immense therapeutic potential. The integration of foundational knowledge with advanced methodological tools is crucial for decrypting the complexities of the linear ubiquitin code. Future research must focus on developing even more specific chemical probes and drugs to target LUBAC and its associated deubiquitinases. Furthermore, exploring the role of M1-chains in specific disease contexts, such as sepsis-induced organ damage and cancer immunotherapy, and understanding its interplay with other ubiquitin linkages through branched chains represent the next frontier. For drug development professionals, the linear ubiquitin pathway presents a promising, albeit complex, target for a new class of anti-inflammatory and immuno-oncology therapeutics, urging a deeper investment in translational research.
References
- 1. LUBAC-mediated linear ubiquitination: a crucial regulator of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8019854/]
- 2. Mechanisms underlying linear ubiquitination and implications ... [https://biosignaling.biomedcentral.com/articles/10.1186/s12964-023-01239-5]
- 3. The linear ubiquitin chain assembly complex (LUBAC) ... [https://elifesciences.org/articles/60660]
- 4. The mechanism of linear ubiquitination in regulating cell ... [https://www.nature.com/articles/s41419-023-06183-3]
- 5. Linear Ubiquitin Chains: Cellular Functions and Strategies ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6966713/]
- 6. Structural Insights into SHARPIN-Mediated Activation of ... [https://www.sciencedirect.com/science/article/pii/S2211124717313062]
- 7. Cooperative Domain Formation by Homologous Motifs in ... [https://pubmed.ncbi.nlm.nih.gov/29694895/]
- 8. Synergistic involvement of the NZF domains of the LUBAC ... [https://www.nature.com/articles/s41419-024-07199-z]
- 9. LUBAC modulates CBM complex functions downstream of ... [https://www.nature.com/articles/s41467-025-65879-6]
- 10. Review Ubiquitin—A structural perspective [https://www.sciencedirect.com/science/article/abs/pii/S1097276524010104]
- 11. Review OTULIN in NF-κB signaling, cell death, and disease [https://www.sciencedirect.com/science/article/abs/pii/S1471490621000983]
- 12. CYLD, A20 and OTULIN deubiquitinases in NF-κB ... [https://www.nature.com/articles/cdd201746]
- 13. independent functions of OTULIN in cell fate control and ... [https://www.nature.com/articles/s41418-020-00675-x]
- 14. OTULIN restricts Met1-linked ubiquitination to control innate ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4194427/]
- 15. CYLD, A20 and OTULIN deubiquitinases in NF-κB ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5520167/]
- 16. CYLD in health and disease - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC10320722/]
- 17. SPATA2 restricts OTULIN-dependent LUBAC activity ... [https://www.sciencedirect.com/science/article/pii/S2211124722018654]
- 18. M1‐linked ubiquitination facilitates NF‐κB activation and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9543601/]
- 19. Linear ubiquitination-mediated NF-κB regulation and its related disorders - PubMed [https://pubmed.ncbi.nlm.nih.gov/23969028/]
- 20. Monitoring the Levels of Cellular NF-κB Activation States [https://pmc.ncbi.nlm.nih.gov/articles/PMC8582385/]
- 21. NF-κB signaling in inflammation [https://www.nature.com/articles/sigtrans201723]
- 22. Exploring TNFR1: from discovery to targeted therapy development | Journal of Translational Medicine | Full Text [https://translational-medicine.biomedcentral.com/articles/10.1186/s12967-025-06122-0]
- 23. A physical wiring diagram for the human immune system [https://www.nature.com/articles/s41586-022-05028-x]
- 24. Researchers discover new way to study protein, could lead ... [https://news.ohsu.edu/2025/03/31/researchers-discover-new-way-to-study-protein-could-lead-to-new-treatments-for-immune-system-diseases]
- 25. M1-linked ubiquitination by LUBEL is required for ... [https://www.nature.com/articles/s41418-018-0164-x]
- 26. Lys63/Met1-hybrid ubiquitin chains are commonly formed ... [https://www.sciencedirect.com/science/article/pii/S0006291X16306647]
- 27. Ubiquitin signaling in immune responses - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC4822134/]
- 28. Hypoxia-induced mitoROS triggers M1 linear ubiquitin ... [https://pubmed.ncbi.nlm.nih.gov/41278941/]
- 29. M1‐linked ubiquitination facilitates NF‐κB activation and ... [https://research.abo.fi/en/publications/m1linked-ubiquitination-facilitates-nf%CE%BAb-activation-and-survival-/]
- 30. Neuro-immune crosstalk in cancer: mechanisms and ... [https://www.nature.com/articles/s41392-025-02241-8]
- 31. STING induces HOIP-mediated synthesis of M1 ubiquitin ... [https://pubmed.ncbi.nlm.nih.gov/39578541/]
- 32. Ubiquitination in macrophage plasticity: from inflammatory to ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12507854/]
- 33. Ubiquitination in macrophage plasticity: from inflammatory ... [https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2025.1621328/full]
- 34. Ubiquitylation of MFHAS1 by the ubiquitin ligase praja2 ... [https://www.nature.com/articles/cddis2017102]
- 35. Inflammation: a matter of immune cell life and death [https://www.nature.com/articles/s44385-025-00010-4]
- 36. Cell death signaling and immune regulation - PubMed Central [https://pmc.ncbi.nlm.nih.gov/articles/PMC12355773/]
- 37. Data-independent acquisition method for ubiquitinome ... [https://www.nature.com/articles/s41467-020-20509-1]
- 38. Linear Ubiquitin Chains: Cellular Functions and Strategies ... [https://www.frontiersin.org/journals/chemistry/articles/10.3389/fchem.2019.00915/full]
- 39. M1-linked ubiquitination facilitates NF-κB activation and ... [https://pubmed.ncbi.nlm.nih.gov/35263507/]
- 40. Mass Spectrometry Proteomics using TUBEs [https://lifesensors.com/mass-spectromtry-proteomics-using-tubes/]
- 41. Tandem Ubiquitin Binding Entities (TUBEs) [https://lifesensors.com/our-technologies/tandem-ubiquitin-binding-entities-tubes/]
- 42. Unraveling chain specific ubiquitination in cells using tandem ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12217038/]
- 43. Unraveling chain specific ubiquitination in cells using ... [https://www.nature.com/articles/s41598-025-07242-9]
- 44. Systematic HOIP interactome profiling reveals critical roles ... [https://www.nature.com/articles/s41467-024-47289-2]
- 45. Current methodologies in protein ubiquitination ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9373315/]
- 46. Protein Ubiquitination: Assays, Sites & Proteomics Methods [https://www.creative-proteomics.com/resource/protein-ubiquitination-detection-assays-site-identification-proteomics.htm]
- 47. Met1-linked ubiquitin signalling in health and disease [https://pmc.ncbi.nlm.nih.gov/articles/PMC7862443/]
- 48. The linear ubiquitin assembly complex (LUBAC) is essential ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4076580/]
- 49. A Ubiquitin Replacement Strategy in Human Cells Reveals ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2779160/]
- 50. Structural basis of K11/K48-branched ubiquitin chain ... [https://www.nature.com/articles/s41467-025-64719-x]
- 51. An NF-kB/TNF-alpha signalling feedback loop acts to ... [https://www.nature.com/articles/s41536-025-00414-1]
- 52. Molecular mechanism of NF-κB-mediated transcriptional ... [https://pubmed.ncbi.nlm.nih.gov/41274683/?utm_source=FeedFetcher&utm_medium=rss&utm_campaign=None&utm_content=0nK-FGmcPzruPpJferMpyd8mRld7hsB1ra05TRUDXN6&fc=None&ff=20251123021540&v=2.18.0.post22+67771e2]
- 53. NF-kB oscillation profiles decode response to anti-EGFR ... [https://www.sciencedirect.com/science/article/pii/S2472555225000127]
- 54. Assessing Immune Function by Profiling Cytokine Release ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3183136/]
- 55. Dictionary of immune responses to cytokines at single-cell ... [https://www.nature.com/articles/s41586-023-06816-9]
- 56. Linear ubiquitination signals in adaptive immune responses [https://pmc.ncbi.nlm.nih.gov/articles/PMC4506786/]
- 57. LUBAC enables tumor-promoting LTβ receptor signaling ... [https://www.nature.com/articles/s41418-024-01355-w]
- 58. OTULIN enzyme found to control tau production in ... [https://www.drugtargetreview.com/news/190936/otulin-enzyme-found-to-control-tau-production-in-alzheimers/]
- 59. Novel Discovery Reveals How Brain Protein OTULIN ... [https://www.wric.com/business/press-releases/ein-presswire/869876629/novel-discovery-reveals-how-brain-protein-otulin-controls-tau-expression-and-could-transform-alzheimers-treatment]
- 60. Molecular bases for HOIPINs-mediated inhibition of LUBAC ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7125101/]
- 61. RNA-Seq reveals how brain protein OTULIN controls tau ... [https://www.rna-seqblog.com/rna-seq-reveals-how-brain-protein-otulin-controls-tau-expression-and-could-transform-alzheimers-treatment/]
- 62. OTULIN orchestrates NCOA4-FTH1 complex to alleviate ... [https://www.sciencedirect.com/science/article/abs/pii/S1567576925004801]
- 63. Signaling to survive: linear ubiquitination in health and ... [https://link.springer.com/article/10.1007/s12268-025-2402-3]
- 64. Ubiquitylomics: An Emerging Approach for Profiling Protein ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11634490/]
- 65. Linear ubiquitination induces NEMO phase separation to ... [https://www.life-science-alliance.org/content/6/4/e202201607]
- 66. Mitochondrial respiratory chain deficiency is associated ... [https://www.nature.com/articles/s41514-025-00273-6]
- 67. Exosome circ-CBLB promotes M1 macrophage ... [https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2025.1627389/full]
- 68. Identification of an M1 Macrophages‐Related Signature for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12513507/]
- 69. Emerging functions of branched ubiquitin chains [https://www.nature.com/articles/s41421-020-00237-y]
- 70. at a glance Linear ubiquitination [https://pubmed.ncbi.nlm.nih.gov/30659056/]
- 71. Activation of the canonical IKK complex by K63/M1-linked ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3780889/]
- 72. LUBAC modulates CBM complex functions downstream of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12602705/]
- 73. LUBAC deficiency perturbs TLR3 signaling to cause ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5110014/]
- 74. Linear ubiquitination: a newly discovered regulator of cell ... [https://www.sciencedirect.com/science/article/abs/pii/S0968000412001909]
- 75. The Role of Protein Ubiquitination in the Onset and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12248740/]
- 76. Extracting Better Ubiquitin Data from your Samples [https://bitesizebio.com/31971/extracting-better-ubiquitin-data-samples/]
- 77. (10X) | Cell Signaling Technology Lysis Buffer Cell [https://www.cellsignal.com/products/buffers-dyes/cell-lysis-buffer-10x/9803]
- 78. For DNA Extraction: Importance, Recipe and Preparation Lysis Buffer [https://geneticeducation.co.in/lysis-buffer-for-dna-extraction/]
- 79. | Thermo Fisher Scientific - US Cell Lysis Buffers [https://www.thermofisher.com/us/en/home/life-science/protein-biology/protein-purification-isolation/cell-lysis-fractionation/cell-lysis-total-protein-extraction.html]
- 80. diGLY proteomics as an approach to identify and quantify the... Ubiquitin [https://pmc.ncbi.nlm.nih.gov/articles/PMC6791129/]
- 81. Ubiquitin signaling in immune responses | Cell Research [https://www.nature.com/articles/cr201640]
- 82. Mechanisms underlying linear ubiquitination and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10685518/]
- 83. Mixed-linkage ubiquitin chains send mixed messages - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3654000/]
- 84. The K48-K63 Branched Ubiquitin Chain Regulates NF-κB ... [https://www.sciencedirect.com/science/article/pii/S1097276516305639]
- 85. Emerging functions of branched ubiquitin chains - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC7835216/]
- 86. Structural basis of K11/K48- branched recognition by... ubiquitin chain [https://www.nature.com/articles/s41467-025-64719-x?error=cookies_not_supported&code=83fe6a1c-ff6c-4d51-a672-4c600e1d8cea]
- 87. K48- and K -linked 63 interactome reveals ubiquitin - and... chain branch [https://pmc.ncbi.nlm.nih.gov/articles/PMC11109483/]
- 88. Assembly and Function of Heterotypic Ubiquitin Chains in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5669814/]
- 89. B cells induced regulatory T cells attenuated the classical ... [https://www.nature.com/articles/s41598-025-19445-1]
- 90. M1 Macrophages Are More Susceptible to Necroptosis [https://www.scientificarchives.com/article/m1-macrophages-are-more-susceptible-to-necroptosis]
- 91. Quantitative characterization and analysis of the dynamic NF ... [https://bmcbioinformatics.biomedcentral.com/articles/10.1186/1471-2105-12-276]
- 92. Ubiquitination-activated TAB–TAK1–IKK–NF-κB axis ... [https://elifesciences.org/reviewed-preprints/106901/reviews]
- 93. In vitro benchmarking of NF-κB inhibitors - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC8867604/]
- 94. In vitro benchmarking of NF-κB inhibitors [https://www.sciencedirect.com/science/article/abs/pii/S001429992030073X]
- 95. Linear Ubiquitination of Hemocyanin Mediated by LUBEL ... [https://www.mdpi.com/1422-0067/26/11/5110]